Химическое строение белков. Строение белков
Белки являются сложными органическими соединениями, состоящими из аминокислот. Химический анализ показал, что белки состоят из следующих элементов:
Углерод 50-55 %
Водород 6-7 %
Кислород 21-23 %
Азот 15-17 %
Сера 0,3-2,5 %.
В составе отдельных белков обнаружены также фосфор, йод, железо, медь и др. макро- и микровещества.
Содержание основных химических элементов может различаться в отдельных белках, исключение составляет азот, среднее количество которого характеризуется наибольшим постоянством и составляет 16 %. В связи с этим существует способ определения количества белка по входящему в его состав азоту. Зная, что 6,25 грамм белка содержит 1 грамм азота, можно найти количество белка, умножив найденное количество азота на коэффициент 6,25.
2. 4. Аминокислоты.
Аминокислоты – карбоновые кислоты альфа-углеродный атом водорода которых замещен на аминогруппу. Белки состоят из аминокислот. В настоящее время известно более 200 различных аминокислот. В организме человека их около 60, а в состав белков входят только 20 аминокислот, которые называют природными или протеиногенными. 19 из них являются альфа-аминокислотами, это означает, что аминогруппа присоединена к альфа-углеродному атому карбоновой кислоты. Общая формула этих аминокислот выглядит следующим образом.
Только аминокислота пролин не соответствует этой формуле, её относят к иминокислотам.
Химические названия аминокислот, для краткости сокращают, например, глутаминовая кислота ГЛУ, серин СЕР и т.д. для записи первичной структуры белков в последнее время стали пользоваться только однобуквенными символами.
Во всех аминокислотах есть общие группировки: -СН2, -NН2, -СООН, они придают общие химические свойства белкам, и радикалы, химическая природа которых разнообразна. Именно они определяют структурные и функциональные особенности аминокислот.
Классификации аминокислот основана на их физико-химических свойствах.
По строению радикалов:
Циклические - гомоциклические ФЕН, ТИР, гетероциклические ТРИ, ГИС.
Ациклические – моноаминомонокарбоновые ГЛИ, АЛА, СЕР, ЦИС, ТРЕ, МЕТ, ВАЛ, ЛЕЙ, ИЛЕЙ,НЛЕЙ, моноаминодикарбоновые АСП, ГЛУ, диаминомонокарбоновые ЛИЗ, АРГ.
По образованию в организме:
Заменимые – могут синтезироваться в организме из веществ белковой и небелковой природы.
Незаменимые – не могут синтезироваться в организме, поэтому должны поступать только с пищей – все циклические аминокислоты, ТРЕ, ВАЛ, ЛЕЙ, ИЛЕЙ.
Биологическое значение аминокислот:
Входят в состав белков организма человека.
Входят в состав пептидов организма человека.
Из аминокислот образованы в организме многие низкомолекулярные биологически активные вещества: ГАМК, биогенные амины и т.д.
Часть гормонов в организме – производные аминокислот (гормоны щитовидной железы, адреналин).
Предшественники азотистых оснований, входящих в состав нуклеиновых кислот.
Предшественники порфиринов, идущих на биосинтез гема для гемоглобина и миоглобина.
Предшественники азотистых оснований, входящих в состав сложных липидов (холина, этаноламина).
Участвуют в биосинтезе медиаторов в нервной системе (ацетилхолин, дофамин, серотонин, норадреналин и др.).
Свойства аминокислот:
Хорошо растворимы в воде.
В водном растворе существуют в виде равновесной смеси биполярного иона, катионной и анионной форм молекулы. Равновесие зависит от рН среды.
NH3-CH-COOH NH3-CH-COO NH2-CH-COO
R + ОН R R + Н
Катионная форма Биполярный ион Анионная форма
Щелочная среда рН Кислая среда
Способны двигаться в электрическом поле, что используется для разделения аминокислот с помощью электрофореза.
Проявляют амфотерные свойства.
Могут играть роль буферной системы, т.к. могут реагировать как слабое основание и слабая кислота.
Белки являются биополимерами, мономерами которых являются остатки альфа-аминокислот, соединенные между собой посредством пептидных связей. Аминокислотная последовательность каждого белка строго определена, в живых организмах она зашифрована посредством генетического кода, на основе считывания которого и происходит биосинтез белковых молекул. В построении белков участвует 20 аминокислот.
Различают следующие виды структуры белковых молекул:
- Первичная. Представляет собой аминокислотную последовательность в линейной цепи.
- Вторичная. Это более компактная укладка полипептидных цепей при помощи формирования водородных связей между пептидными группами. Есть два варианта вторичной структуры – альфа-спираль и бета-складчатость.
- Третичная. Представляет собой укладку полипептидной цепочки в глобулу. При этом формируются водородные, дисульфидные связи, также стабилизация молекулы реализуется благодаря гидрофобным и ионным взаимодействиям аминокислотных остатков.
- Четвертичная. Белок состоит из нескольких полипептидных цепей, которые взаимодействуют между собой посредством нековалентных связей.
Таким образом, соединенные в определенной последовательности аминокислоты образуют полипептидную цепь, отдельные части которой сворачиваются в спираль или формируют складки. Такие элементы вторичной структур образуют глобулы, формируя третичную структуру белка. Отдельные глобулы взаимодействуют между собой, образуя сложные белковые комплексы с четвертичной структурой.
Классификация белков
Существует несколько критериев, по которым можно классифицировать белковые соединения. По составу различают простые и сложные белки. Сложные белковые вещества содержат в своем составе неаминокислотные группы, химическая природа которых может быть различной. В зависимости от этого выделяют:
- гликопротеины;
- липопротеины;
- нуклеопротеины;
- металлопротеиды;
- фосфопротеины;
- хромопротеиды.
Также существует классификация по общему типу строения:
- фибриллярные;
- глобулярные;
- мембранные.
Протеинами называют простые (однокомпонентные) белки, состоящие лишь из остатков аминокислот. В зависимости от растворимости они делятся на следующие группы:
Подобная классификация не совсем точна, ведь согласно последним исследованиям многие простые белки связаны с минимальным количеством небелковых соединений. Так, в состав некоторых протеинов входят пигменты, углеводы, иногда липиды, что делает их больше похожими на сложные белковые молекулы.
Физико-химические свойства белка
Физико-химические свойства белков обусловлены составом и количеством входящих в их молекулы остатков аминокислот. Молекулярные массы полипептидов сильно колеблются: от нескольких тысяч до миллиона и более. Химические свойства белковых молекул разнообразны, включают в себя амфотерность, растворимость, а также способность к денатурации.
Амфотерность
Поскольку в состав белков входят и кислые, и основные аминокислоты, то всегда в составе молекулы будут свободные кислые и свободные основные группы (СОО- и NН3+ соответственно). Заряд определяется соотношением основных и кислых аминокислотных групп. По этой причине белки заряжены “+”, если уменьшается рН, и наоборот, “-”, если рН увеличивается. В случае, когда рН соответствует изоэлектрической точке, белковая молекула будет иметь нулевой заряд. Амфотерность важна для осуществления биологических функций, одной из которых является поддержание уровня рН в крови.
Растворимость
Классификация белков по свойству растворимости уже была приведена выше. Растворимость белковых веществ в воде объясняется двумя факторами:
- заряд и взаимное отталкивание белковых молекул;
- формирование гидратной оболочки вокруг белка – диполи воды взаимодействуют с заряженными группами на внешней части глобулы.
Денатурация
Физико-химическое свойство денатурации представляет собой процесс разрушения вторичной, третичной структуры белковой молекулы под влиянием ряда факторов: температуры, действии спиртов, солей тяжелых металлов, кислот и других химических агентов.
Важно! Первичная структура при денатурации не разрушается.
Химические свойства белков, качественные реакции, уравнения реакций
Химические свойства белков можно рассмотреть на примере реакций их качественного обнаружения. Качественные реакции позволяют определить наличие пептидной группы в соединении:
1. Ксантопротеиновая. При действии на белок азотной кислоты высокой концентрации образуется осадок, который при нагревании приобретает желтый цвет.
2. Биуретовая. При действии на слабощелочной раствор белка сульфата меди образуются комплексные соединения между ионами меди и полипептидами, что сопровождается окрашиванием раствора в фиолетово-синий цвет. Реакция используется в клинической практике для определения концентрации белка в сыворотке крови и других биологических жидкостях.
Еще одним важнейшим химическим свойством является обнаружение серы в белковых соединениях. С этой целью щелочной раствор белка нагревают с солями свинца. При этом получают черный осадок, содержащий сульфид свинца.
Биологическое значение белка
Благодаря своим физическим и химическим свойствам белки выполняют большое количество биологических функций, в перечень которых входят:
- каталитическая (белки-ферменты);
- транспортная (гемоглобин);
- структурная (кератин, эластин);
- сократительная (актин, миозин);
- защитная (иммуноглобулины);
- сигнальная (рецепторные молекулы);
- гормональная (инсулин);
- энергетическая.
Белки важны для организма человека, поскольку участвуют в образовании клеток, обеспечивают сокращение мышц у животных, вместе с сывороткой крови переносят многие химические соединения. Помимо этого, белковые молекулы являются источником незаменимых аминокислот и осуществляют защитную функцию, участвуя в выработке антител и формировании иммунитета.
ТОП-10 малоизвестных фактов о белке
- Белки начали изучать с 1728 года, именно тогда итальянец Якопо Бартоломео Беккари выделил белок из муки.
- Сейчас широкое распространение получили рекомбинантные белки. Их синтезируют путем модификации генома бактерий. В частности, таким способом получают инсулин, факторы роста и другие белковые соединения, которые используют в медицине.
- У антарктических рыб были обнаружены белковые молекулы, предотвращающие замерзание крови.
- Белок резилин отличается идеальной эластичностью и является основой мест крепления крыльев насекомых.
- В организме есть уникальные белки шапероны, которые способны восстанавливать корректную нативную третичную или четвертичную структуру других белковых соединений.
- В ядре клетки присутствуют гистоны – белки, которые принимают участие в компактизации хроматина.
- Молекулярную природу антител – особых защитных белков (иммуноглобулинов) – начали активно изучать с 1937 года. Тиселиус и Кабат применяли электрофорез и доказали, что у иммунизированных животных увеличена гамма-фракция, а после абсорбции сыворотки провоцирующим антигеном, распределение белков по фракциям возвращалось к картине интактного животного.
- Яичный белок – яркий пример реализации белковыми молекулами резервной функции.
- В молекуле коллагена каждый третий аминокислотный остаток образован глицином.
- В составе гликопротеинов 15-20% составляют углеводы, а в составе протеогликанов их доля – 80-85%.
Заключение
Белки – сложнейшие соединения, без которых сложно представить жизнедеятельность любого организма. Выделено более 5000 белковых молекул, но каждый индивидуум обладает собственным набором белков и этим отличается от других особей своего вида.
Важнейшие химические и физические свойства белков обновлено: Март 21, 2019 автором: Научные Статьи.Ру
Белки представляют собой высокомолекулярные органические соединения, построенные из остатков 20 аминокислот. По своей структуре они относятся к полимерам. Их молекулы имеют форму длинных цепей, состоящих из повторяющихся молекул – мономеров. Для образования полимерной молекулы каждый из мономеров должен обладать как минимум двумя реакционноспособными связями с другими мономерами.
Белок по своей структуре похож на полимер найлон: оба полимера представляют собой цепочку мономеров. Но между ними есть существенное различие. Найлон состоит из двух видов мономеров, а белок построен из 20 различных мономеров – аминокислот. В зависимости от порядка чередования мономеров образуется множество различных видов белков.
Общая формула аминокислот, образующих белок, имеет вид:
Из данной формулы видно, что к центральному атому углерода присоединены четыре разные группы. Три из них – атом водорода Н, щелочная аминогруппа Н N и карбоксильная группа СООН – для всех аминокислот одинаковы. По составу и структуре четвертой группы, обозначенной R , аминокислоты отличаются друг от друга. В самых простых случаях в молекуле глицерина – такая группа представляет собой атом водорода, в молекуле аланина – СН и т. д.
Химическая связь (– СО – NH –), соединяющая аминогруппу одной аминокислоты с карбоксильной группой другой в молекулах белков, называется пептидной связью (см. рис.7.5).

Все активные организмы, будь то растения, животные, бактерии или вирусы, содержат белки, построенные из одних и тех же аминокислот. Поэтому в любом виде пищи содержатся те же аминокислоты, которые входят в состав белков организмов, потребляющих пищу.
В определении «белки – это полимеры, построенные из 20 разных аминокислот» содержится неполная характеристика белков. В лабораторных условиях не составляет труда в растворе аминокислот получить пептидные связи и сформировать таким образом длинные молекулярные цепи. Однако в таких цепях расположение аминокислот будет хаотическим, и образовавшиеся молекулы будут отличаться друг от друга. В то же время в каждом из природных белков порядок расположения отдельных видов аминокислот всегда один и тот же. А это означает, что при синтезе белка в живой системе используется информация, в соответствии с которой формируется вполне определенная последовательность аминокислот для каждого белка.
Последовательность расположения аминокислот в белке определяет его пространственную структуру. Большинство белков выполняют функцию катализаторов. Вих пространственной структуре есть активные центры в виде углублений с вполне определенной формой. В такие центры попадают молекулы, превращение которых катализируется данным белком. Белок, выступающий в данном случае в роли фермента, может катализировать реакцию только при совпадении по форме превращающейся молекулы и активного центра. Этим и определяется высокая селективность белка-фермента.
Активный центр фермента может образовываться в результате свертывания весьма удаленных друг от друга участков белковой цепи. Поэтому замена одной аминокислоты другой даже на небольшом расстоянии от активного центра может повлиять на селективность фермента, либо полностью разрушить центр. Создавая различные последовательности аминокислот, можно получать самые разнообразные активные центры. В этом заключается одна из важнейших особенностей белков, выступающих в роли ферментов.
Среди органических веществ белки , или протеины , - самые многочисленные, наиболее разнообразные и имеющие первостепенное значение биополимеры. На их долю приходится 50 - 80% сухой массы клетки.
Молекулы белков имеют большие размеры, поэтому их называют макромолекулами . Кроме углерода , кислорода , водорода и азота , в состав белков могут входить сера, фосфор и железо. Белки отличаются друг от друга числом (от ста до нескольких тысяч), составом и последовательностью мономеров. Мономерами белков являются аминокислоты (рис. 1)
Бесконечное разнообразие белков создается за счет различного сочетания всего 20 аминокислот. Каждая аминокислота имеет свое название, особое строение и свойства. Их общую формулу можно представить в следующем виде:
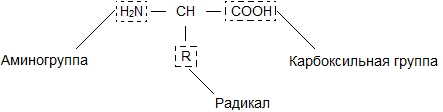
Молекула аминокислоты состоит из двух одинаковых для всех аминокислот частей, одна из которых является аминогруппой (-NH 2 ) с основными свойствами, другая - карбоксильной группой (-COOH ) с кислотными свойствами. Часть молекулы, называемая радикалом (R ), у разных аминокислот имеет различное строение. Наличие в одной молекуле аминокислоты основной и кислотной групп обусловливает их высокую реакционную способность. через эти группы происходит соединение аминокислот при образовании белка. При этом возникает молекула воды, а освободившиеся электроны образуют пептидную связь. Поэтому белки называют полипептидами .
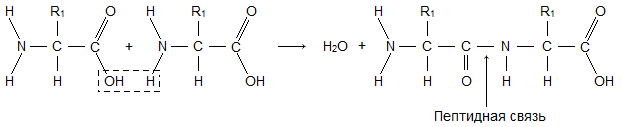
Молекулы белков могут иметь различные пространственные конфигурации, и в их строении различают четыре уровня структурной организации.
Последовательность аминокислот в составе полипептидной цепи представляет первичную структуру
белка. Она уникальна для любого белка и определяет его форму, свойства и функции.
Большинство белков имеют вид спирали в результате образования водородных связей между -CO-
и -NH-
группами разных аминокислотных остатков полипептидной цепи. Водородные связи малопрочные, но в комплексе они обеспечивают довольно прочную структуру. Эта спираль - вторичная структура
белка.
Третичная структура - трехмерная пространственная «упаковка» полипептидной цепи. В результате возникает причудливая, но для каждого белка специфическая конфигурация - глобула . Прочность третичной структуры обеспечивается разнообразными связями, возникающими между радикалами аминокислот.
Четвертичная структура
характерна не для всех белков. Она возникает в результате соединения нескольких макромолекул с третичной структурой в сложный комплекс. Например, гемоглобин крови человека представляет комплекс из четырех макромолекул белка.
Такая сложность структуры белковых молекул связана с разнообразием функций, свойственных этим биополимерам.
Нарушение природной структуры белка называют денатурацией
. Она может происходить под воздействием температуры, химических веществ, лучистой энергии и других факторов. При слабом воздействии распадается только четвертичная структура, при более сильном - третичная, а затем - вторичная, и белок остается в виде полипептидной цепи.
Этот процесс частично обратим: если не нарушена первичная структура, то денатурированный белок способен восстанавливать свою структуру. Отсюда следует, что все особенность строение макромолекулы белка определяются его первичной структурой.
Кроме простых белков , состоящих только из аминокислот, есть еще и сложные белки
В организмах живых существ содержится более 100 различных аминокислот, однако, в строительстве белков используются не все, а только 20, так называемых «фундаментальных». В табл. 1 приведены их названия (большинство названий сложилось исторически), структурная формула, а также широко применяемое сокращенное обозначение. Все структурные формулы расположены в таблице таким образом, чтобы основной
фрагмент аминокислоты находился справа.| Таблица 1. АМИНОКИСЛОТЫ, УЧАСТВУЮЩИЕ В СОЗДАНИИ БЕЛКОВ . | ||
| Название | Структура |
Обозначение |
| ГЛИЦИН |  |
ГЛИ |
| АЛАНИН |  |
АЛА |
| ВАЛИН |  |
ВАЛ |
| ЛЕЙЦИН |  |
ЛЕЙ |
| ИЗОЛЕЙЦИН |  |
ИЛЕ |
| СЕРИН |  |
СЕР |
| ТРЕОНИН |  |
ТРЕ |
| ЦИСТЕИН |  |
ЦИС |
| МЕТИОНИН |  |
МЕТ |
| ЛИЗИН |  |
ЛИЗ |
| АРГИНИН |  |
АРГ |
| АСПАРАГИНОВАЯ КИСЛОТА |  |
АСП |
| АСПАРАГИН |  |
АСН |
| ГЛУТАМИНОВАЯ КИСЛОТА |  |
ГЛУ |
| ГЛУТАМИН |  |
ГЛН |
| ФЕНИЛАЛАНИН |  |
ФЕН |
| ТИРОЗИН |  |
ТИР |
| ТРИПТОФАН |  |
ТРИ |
| ГИСТИДИН |  |
ГИС |
| ПРОЛИН |  |
ПРО |
| В международной практике принято сокращенное обозначение перечисленных аминокислот с помощью латинских трехбуквенных или однобуквенных сокращений, например, глицин – Gly или G, аланин – Ala или A. | ||
Среди этих двадцати аминокислот (табл. 1) только пролин содержит рядом с карбоксильной группой СООН группу
NH (вместо NH 2), так как она входит в состав циклического фрагмента.Восемь аминокислот (валин, лейцин, изолейцин, треонин, метионин, лизин, фенилаланин и триптофан), помещенные в таблице на сером фоне, называют незаменимыми, поскольку организм для нормального роста и развития должен постоянно получать их с белковой пищей.
Белковая молекула образуется в результате последовательного соединения аминокислот, при этом карбоксильная группа одной кислоты взаимодействует с аминогруппой соседней молекулы, в результате образуется пептидная связь
CO NH и выделяется молекула воды. На рис. 1 показано последовательное соединение аланина, валина и глицина.
Рис.1. ПОСЛЕДОВАТЕЛЬНОЕ СОЕДИНЕНИЕ АМИНОКИСЛОТ при образовании белковой молекулы. В качестве основного направления полимерной цепи выбран путь от концевой аминогруппы
H 2 N к концевой карбоксильной группе COOH .Чтобы компактно описать строение белковой молекулы, используют сокращенные обозначения аминокислот (табл. 1, третий столбец), участвующих в образовании полимерной цепи. Фрагмент молекулы, показанный на рис. 1, записывают следующим образом:
H 2 N -АЛА-ВАЛ-ГЛИ- COOH .Белковые молекулы содержат от 50 до 1500 аминокислотных остатков (более короткие цепи называют полипептидами). Индивидуальность белка определяется набором аминокислот, из которых составлена полимерная цепь и, что не менее важно, порядком их чередования вдоль цепи. Например, молекула инсулина состоит из 51 аминокислотного остатка (это один из самых короткоцепных белков) и представляет собой две соединенных между собой параллельных цепи неодинаковой длины. Порядок чередования аминокислотных фрагментов показан на рис. 2.
Молекулы аминокислоты цистеина (табл. 1) содержат реакционно-способные сульфгидридные группы
SH , которые взаимодействуют между собой, образуя дисульфидные мостики S - S -. Роль цистеина в мире белков особая, с его участием образуются поперечные сшивки между полимерными белковыми молекулами.Объединение аминокислот в полимерную цепь происходит в живом организме под управлением нуклеиновых кислот, именно они обеспечивают строгий порядок сборки и регулируют фиксированную длину полимерной молекулы (см . НУКЛЕИНОВЫЕ КИСЛОТЫ ).
Структура белков. Состав белковой молекулы, представленный в виде чередующихся остатков аминокислот (рис. 2), называют первичной структурой белка. Между присутствующими в полимерной цепи имино-группами HN и карбонильными группами CO возникают водородные связи (см. ВОДОРОДНАЯ СВЯЗЬ ), в результате молекула белка приобретает определенную пространственную форму, называемую вторичной структурой. Наиболее распространены два типа вторичной структуры белков.Первый вариант, называемый
a -спиралью, реализуется с помощью водородных связей внутри одной полимерной молекулы. Геометрические параметры молекулы, определяемые длинами связей и валентными углами, таковы, что образование водородных связей оказывается возможным для групп H - N и C = O , между которыми находятся два пептидных фрагмента H - N - C = O (рис. 3).Состав полипептидной цепи, показанной на рис. 3, записывают в сокращенном виде следующим образом:
H 2 N -АЛА ВАЛ-АЛА-ЛЕЙ-АЛА-АЛА-АЛА-АЛА-ВАЛ-АЛА-АЛА-АЛА- COOH .В результате стягивающего действия водородных связей молекула приобретает форму спирали так называемая
a -спираль, ее изображают в виде изогнутой спиралевидной ленты, проходящей через атомы, образующие полимерную цепь (рис. 4)Другой вариант вторичной структуры, называемый
b -структурой, образуется также при участии водородных связей, отличие состоит в том, что взаимодействуют группы H - N и C = O двух или более полимерных цепей, расположенных параллельно. Поскольку полипептидная цепь имеет направление (рис. 1), возможны варианты, когда направление цепей совпадает (параллельная b -структура, рис. 5), либо они противоположны (антипараллельная b -структура, рис. 6).В образовании
b -структуры могут участвовать полимерные цепи различного состава, при этом органические группы, обрамляющие полимерную цепь (Ph , CH 2 ОН и др.), в большинстве случаев играют второстепенную роль, решающее значение имеет взаиморасположение групп H - N и C = O . Поскольку относительно полимерной цепи H - N и C = O группы направлены в различные стороны (на рисунке вверх и вниз), становится возможным одновременное взаимодействие трех и более цепей.Состав первой полипептидной цепи на рис. 5:
H 2 N -ЛЕЙ-АЛА-ФЕН-ГЛИ-АЛА-АЛА- COOHСостав второй и третей цепи:
H 2 N -ГЛИ-АЛА-СЕР-ГЛИ-ТРЕ-АЛА- COOHСостав полипептидных цепей, показанных на рис. 6, тот же, что и на рис. 5, отличие в том, что вторая цепь имеет противоположное (в сравнении с рис. 5) направление.
Возможно образование
b -структуры внутри одной молекулы, когда фрагмент цепи на определенном участке оказывается повернутым на 180°, в этом случае две ветви одной молекулы имеют противоположное направление, в результате образуется антипараллельная b -структура (рис. 7).Структура, показанная на рис. 7 в плоском изображении, представлена на рис. 8 в виде объемной модели. Участки
b -структуры принято упрощенно обозначать плоской волнистой лентой, которая проходит через атомы, образующие полимерную цепь.В структуре многих белков чередуются участки
a -спирали и лентообразные b -структуры, а также одиночные полипептидные цепи. Их взаиморасположение и чередование в полимерной цепи называют третичной структурой белка.Способы изображения структуры белков показаны далее на примере растительного белка крамбина. Структурные формулы белков, содержащих часто до сотни аминокислотных фрагментов, сложны, громоздки и трудны для восприятия, поэтому иногда используют упрощенные структурные формулы без символов химических элементов (рис. 9, вариант А), но при этом сохраняют окраску валентных штрихов в соответствии с международными правилами (рис. 4). Формулу при этом представляют не в плоском, а в пространственном изображении, что соответствует реальной структуре молекулы. Такой способ позволяет, например, различить дисульфидные мостики (подобные тем, которые есть в инсулине, рис. 2), фенильные группы в боковом обрамлении цепи и др. Изображение молекул в виде объемных моделей (шарики, соединенные стержнями) несколько более наглядно (рис. 9, вариант Б). Однако оба способа не позволяют показать третичную структуру, поэтому американский биофизик Джейн Ричардсон предложил изображать
a -структуры в виде спирально закрученных лент (см . рис. 4), b -структуры в виде плоских волнистых лент (рис. 8), а соединяющие их одиночные цепи в форме тонких жгутов, каждый тип структуры имеет свою окраску. Сейчас широко применяют такой способ изображения третичной структуры белка (рис. 9, вариант В). Иногда для большей информативности показывают совместно третичную структуру и упрощенную структурную формулу (рис. 9, вариант Г). Есть и модификации способа, предложенного Ричардсоном: a -спирали изображают в виде цилиндров, а b -структуры в форме плоских стрелок, указывающих и направление цепи (рис. 9, вариант Д). Менее распространен способ, при котором всю молекулу изображают в виде жгута, где неодинаковые структуры выделяют различающейся окраской, а дисульфидные мостики показывают в виде желтых перемычек (рис. 9, вариант Д).Наиболее удобен для восприятия вариант В, когда при изображении третичной структуры особенности строения белка (аминокислотные фрагменты, порядок их чередования, водородные связи) не указывают, при этом исходят из того, что все белки содержат «детали», взятые из стандартного набора двадцати аминокислот (табл. 1). Основная задача при изображении третичной структуры показать пространственное расположение и чередование вторичных структур.
Наиболее удобна для восприятия объемная третичная структура (вариант В), освобожденная от деталей структурной формулы.
Белковая молекула, обладающая третичной структурой, как правило, принимает определенную конфигурацию, которую формируют полярные (электростатические) взаимодействия и водородные связи. В результате молекула приобретает форму компактного клубка глобулярные белки (
globules , лат. шарик) , либо нитевидную фибриллярные белки (fibra , лат. волокно).Пример глобулярной структуры белок альбумин, к классу альбуминов относят белок куриного яйца. Полимерная цепь альбумина собрана, основном, из аланина, аспаргиновой кислоты, глицина, и цистеина, чередующихся в определенном порядке. Третичная структура содержит
a -спирали, соединенные одиночными цепями (рис. 10).Пример фибриллярной структуры белок фиброин. Он содержат большое количество остатков глицина, аланина и серина (каждый второй аминокислотный остаток глицин); остатки цистеина, содержащего сульфгидридные группы, отсутствуют. Фиброин основной компонент натурального шелка и паутины, содержит
b -структуры, соединенные одиночными цепями (рис. 11).Возможность образования третичной структуры определенного типа заложена в первичной структуре белка, т.е. определена заранее порядком чередования аминокислотных остатков. Из определенных наборов таких остатков преимущественно возникают
a -спирали (подобных наборов существует достаточно много), другой набор приводит к появлению b -структур, одиночные цепи характеризуются своим составом.Некоторые белковые молекулы, сохраняя третичную структуру, способны объединяться в крупные надмолекулярные агрегаты, при этом их удерживают вместе полярные взаимодействия, а также водородные связи. Такие образования называют четвертичной структурой белка. Например, белок ферритин, состоящий в основной массе из лейцина, глутаминовой кислоты, аспарагиновой кислоты и гиститдина (в феррицине есть в различном количестве все 20 аминокислотных остатков) образует третичную структуру из четырех параллельно уложенных
a -спиралей. При объединении молекул в единый ансамбль (рис. 12) образуется четвертичная структура, в которую может входить до 24 молекул ферритина.Другой пример надмолекулярных образований структура коллагена. Это фибриллярный белок, цепи которого построены в основном из глицина, чередующегося с пролином и лизином. Структура содержит одиночные цепи, тройные
a -спирали, чередующиеся с лентообразными b -структурами, уложенными в виде параллельных пучков (рис. 13). Химические свойства белков. При действии органических растворителей, продуктов жизнедеятельности некоторых бактерий (молочнокислое брожение) или при повышении температуры происходит разрушение вторичных и третичных структур без повреждения его первичной структуры, в результате белок теряет растворимость и утрачивает биологическую активность, этот процесс называют денатурацией, то есть потерей натуральных свойств, например, створаживание кислого молока, свернувшийся белок вареного куриного яйца. При повышенной температуре белки живых организмов (в частности, микроорганизмов) быстро денатурируют. Такие белки не способны участвовать в биологических процессах, в результате микроорганизмы погибают, поэтому кипяченое (или пастеризованное) молоко может дольше сохраняться.Пептидные связи
H - N - C = O , образующие полимерную цепь белковой молекулы, в присутствии кислот или щелочей гидролизуются, при этом происходит разрыв полимерной цепи, что, в конечном итоге, может привести к исходным аминокислотам. Пептидные связи, входящие в состав a -спиралей или b -структур, более устойчивы к гидролизу и различным химическим воздействиям (по сравнению с теми же связями в одиночных цепях). Более деликатную разборку белковой молекулы на составляющие аминокислоты проводят в безводной среде с помощью гидразина H 2 N NH 2 , при этом все аминокислотные фрагменты, кроме последнего, образуют так называемые гидразиды карбоновых кислот, содержащие фрагмент C (O ) HN NH 2 (рис. 14).Подобный анализ может дать информацию об аминокислотном составе того или иного белка, однако важнее знать их последовательность в белковой молекуле. Одна из широко применяемых для этой цели методик действие на полипептидную цепь фенилизотиоцианата (ФИТЦ), который в щелочной среде присоединяется к полипептиду (с того конца, который содержит аминогруппу), а при изменении реакции среды на кислую, отсоединяется от цепи, унося с собой фрагмент одной аминокислоты (рис. 15).
Разработано много специальных методик для подобного анализа, в том числе и такие, которые начинают «разбирать» белковую молекулу на составляющие компоненты, начиная с карбоксильного конца.
Поперечные дисульфидные мостики
S - S (образовавшиеся при взаимодействии остатков цистеина, рис. 2 и 9) расщепляют, превращая их в HS -группы действием различных восстановителей. Действие окислителей (кислорода или перекиси водорода) приводит вновь к образованию дисульфидных мостиков (рис. 16).Для создания дополнительных поперечных сшивок в белках используют реакционную способность амино- и карбоксильных групп. Более доступны для различных взаимодействий аминогруппы, которые находятся в боковом обрамлении цепи фрагменты лизина, аспарагина, лизина, пролина (табл. 1). При взаимодействии таких аминогрупп с формальдегидом идет процесс конденсации и возникают поперечные мостики
NH CH 2 NH (рис.17).Концевые карбоксильные группы белка способны реагировать с комплексными соединениями некоторых поливалентных металлов (чаще применяют соединения хрома), при этом также возникают поперечные сшивки. Оба процесса используются при дублении кожи.
Роль белков в организме разнообразна.Ферменты (fermentatio лат . брожение), другое их название энзимы (en zumh греч . в дрожжах) это белки, обладающие каталитической активностью, они способны увеличивать скорости биохимических процессов в тысячи раз. Под действием ферментов составные компоненты пищи: белки, жиры и углеводы расщепляются до более простых соединений, из которых затем синтезируются новые макромолекулы, необходимые организму определенного типа. Ферменты принимают участие и во многих биохимических процессах синтеза, например, в синтезе белков (одни белки помогают синтезировать другие).
Ферменты не только высокоэффективные катализаторы, но и селективные (направляют реакцию строго в заданном направлении). В их присутствии реакция проходит практически со 100%-ным выходом без образования побочных продуктов и при этом условия протекания мягкие: обычное атмосферное давление и температура живого организма. Для сравнения, синтез аммиака из водорода и азота в присутствии катализатора активированного железа проводят при 400500° С и давлении 30 МПа, выход аммиака 1525% за один цикл. Ферменты считаются непревзойденными катализаторами.
Интенсивное исследование ферментов началось в середине 19 в., сейчас изучено более 2000 различных ферментов, это самый многообразный класс белков.
Названия ферментов составляют следующим образом: к наименованию реагента, с которым взаимодействует фермент, или к названию катализируемой реакции добавляют окончание -аза , например, аргиназа разлагает аргинин (табл.1), декарбоксилаза катализирует декарбоксилирование, т.е. отщепление СО 2 от карбоксильной группы:
® СН + СО 2Часто, для более точного обозначения роли фермента в его названии указывают и объект, и тип реакции, например, алкогольдегидрогеназа фермент, осуществляющий дегидрирование спиртов.
Для некоторых ферментов, открытых достаточно давно, сохранилось историческое название (без окончания аза), например, пепсин (
pepsis , греч . пищеварение) и трипсин (thrypsis греч . разжижение), эти ферменты расщепляют белки.Для систематизации ферменты объединяют в крупные классы, в основу классификации положен тип реакции, классы именуют по общему принципу название реакции и окончание аза. Далее перечислены некоторые из таких классов.
Оксидоредуктазы ферменты, катализирующие окислительно-восстановительные реакции. Входящие в этот класс дегидрогеназы осуществляют перенос протона, например алкогольдегидрогеназа (АДГ) окисляет спирты до альдегидов, последующее окисление альдегидов до карбоновых кислот катализируют альдегиддегидрогеназы (АЛДГ). Оба процесса происходят в организме при переработке этанола в уксусную кислоту (рис. 18).

Рис. 18. ДВУХСТАДИЙНОЕ ОКИСЛЕНИЕ ЭТАНОЛА до уксусной кислоты
Наркотическим действием обладает не этанол, а промежуточный продукт ацетальдегид, чем ниже активность фермента АЛДГ, тем медленнее проходит вторая стадия окисление ацетальдегида до уксусной кислоты и тем дольше и сильнее проявляется опьяняющее действие от приема внутрь этанола. Анализ показал, что более чем у 80% представителей желтой расы относительно низкая активность АЛДГ и потому заметно более тяжелая переносимость алкоголя. Причина такой врожденной пониженной активности АЛДГ состоит в том, что часть остатков глутаминовой кислоты в молекуле «ослабленной» АЛДГ заменена фрагментами лизина (табл. 1).
Трансферазы ферменты, катализирующие перенос функциональных групп, например, трансиминаза катализирует перемещение аминогруппы.
Гидролазы ферменты, катализирующие гидролиз. Упомянутые ранее трипсин и пепсин осуществляют гидролиз пептидных связей, а липазы расщепляют сложноэфирную связь в жирах:
R С(О)О R 1 +Н 2 О ® R С(О)ОН + НО R 1Лиазы ферменты, катализирующие реакции, которые проходят не гидролитическим путем, в результате таких реакций происходит разрыв связей С-С, С-О, С-N и образование новых связей. Фермент декарбоксилаза относится к этому классу
Изомеразы ферменты, катализирующие изомеризацию, например, превращение малеиновой кислоты в фумаровую (рис. 19), это пример цис транс изомеризации (см . ИЗОМЕРИЯ).

Рис. 19. ИЗОМЕРИЗАЦИЯ МАЛЕИНОВОЙ КИСЛОТЫ в фумаровую в присутствии фермента.
В работе ферментов соблюдается общий принцип, в соответствии с которым всегда есть структурное соответствие фермента и реагента ускоряемой реакции. По образному выражению одного из основателей учения о ферментах Э.Фишера, реагент подходит к ферменту, как ключ к замку. В связи с этим каждый фермент катализирует определенную химическую реакцию или группу реакций одного типа. Иногда фермент может действовать на одно единственное соединение, например, уреаза (
uron греч. моча) катализирует только гидролиз мочевины: (H 2 N) 2 C = O + H 2 O = CO 2 + 2NH 3 Наиболее тонкую избирательность проявляют ферменты, различающие оптически активные антиподы лево- и правовращающие изомеры (см . ИЗОМЕРИЯ ). L -аргиназа действует только на левовращающий аргинин и не затрагивает правовращающий изомер. L-лактатдегидрогеназа действует только на левовращающие эфиры молочной кислоты, так называемые лактаты (lactis лат. молоко), в то время как D-лактатдегидрогеназа расщепляет исключительно D-лактаты.Большая часть ферментов действует не на одно, а на группу родственных соединений, например, трипсин «предпочитает» расщеплять пептидные связи образованные лизином и аргинином (табл. 1.)
Каталитические свойства некоторых ферментов, таких как гидролазы, определяются исключительно строением самой белковой молекулы, другой класс ферментов оксидоредуктазы (например, алкогольдегидрогеназа) могут проявлять активность только в присутствии связанных с ними небелковых молекул витаминов, активирующих ионов Mg, Са,
Zn , Мn и фрагментов нуклеиновых кислот (рис. 20).Транспортные белки связывают и переносят различные молекулы или ионы через мембраны клеток (как внутрь клетки, так и вовне), а также от одного органа к другому.
Например, гемоглобин связывает кислород при прохождении крови через легкие и доставляет его к различным тканям организма, где кислород высвобождается и затем используется для окисления компонентов пищи, этот процесс служит источником энергии (иногда употребляют термин «сжигание» пищевых продуктов в организме).
Помимо белковой части гемоглобин содержит комплексное соединение железа с циклической молекулой порфирином (
porphyros греч. пурпур), что и обусловливает красный цвет крови. Именно этот комплекс (рис. 21, слева) играет роль переносчика кислорода. В гемоглобине порфириновый комплекс железа располагается внутри белковой молекулы и удерживается с помощью полярных взаимодействий, а также координационной связи с азотом в гистидине (табл. 1), входящем в состав белка. Молекула О 2 , которую переносит гемоглобин, присоединяется с помощью координационной связи к атому железа со стороны, противоположной той, к которой присоединен гистидин (рис. 21, справа).Справа показано строение комплекса в форме объемной модели. Комплекс удерживается в белковой молекуле с помощью координационной связи (синий пунктир) между атомом
Fe и атомом N в гистидине, входящим в состав белка. Молекула О 2 , которую переносит гемоглобин, присоединена координационно (красный пунктир) к атому Fe с противоположной страны плоского комплекса.Гемоглобин один из наиболее подробно изученных белков, он состоит из
a -спиралей, соединенных одиночными цепями, и содержит в своем составе четыре комплекса железа. Таким образом, гемоглобин представляет собой как бы объемистую упаковку для переноса сразу четырех молекул кислорода. По форме гемоглобин соответствует глобулярным белкам (рис. 22).Основное «достоинство» гемоглобина состоит в том, что присоединение кислорода и последующее его отщепление при передаче различным тканям и органам проходит быстро. Монооксид углерода, СО (угарный газ), связывается с
Fe в гемоглобине еще быстрее, но, в отличие от О 2 , образует трудно разрушающийся комплекс. В результате такой гемоглобин не способен связывать О 2 , что приводит (при вдыхании больших количеств угарного газа) к гибели организма от удушья.Вторая функция гемоглобина перенос выдыхаемого СО 2 , но в процессе временного связывания углекислоты участвует не атом железа, а
H 2 N -группы белка.«Работоспособность» белков зависит от их строения, например, замена единственного аминокислотного остатка глутаминовой кислоты в полипептидной цепи гемоглобина на остаток валина (изредка наблюдаемая врожденная аномалия) приводит к заболеванию, называемому серповидноклеточная анемия.
Существуют также транспортные белки, способные связывать жиры, глюкозу, аминокислоты и переносить их как внутрь, так и вовне клеток.
Транспортные белки особого типа не переносят сами вещества, а выполняют функции «транспортного регулировщика», пропуская определенные вещества сквозь мембрану (внешнюю стенку клетки). Такие белки чаще называют мембранными. Они имеют форму пустотелого цилиндра и, встраиваясь в стенку мембраны, обеспечивают перемещение некоторых полярных молекул или ионов внутрь клетки. Пример мембранного белка порин (рис. 23).
Пищевые и запасные белки, как следует из названия, служат источниками внутреннего питания, чаще для зародышей растений и животных, а также на ранних стадиях развития молодых организмов. К пищевым белкам относят альбумин (рис. 10) основной компонент яичного белка, а также казеин главный белок молока. Под действием фермента пепсина казеин в желудке створаживается, это обеспечивает его задержку в пищеварительном тракте и эффективное усвоение. Казеин содержит фрагменты всех аминокислот, необходимых организму.
В ферритине (рис. 12), который содержится в тканях животных, запасены ионы железа.
К запасным белкам относят также миоглобин, по составу и строению напоминающий гемоглобин. Миоглобин сосредоточен, главным образом, в мышцах, его основная роль хранение кислорода, который ему отдает гемоглобин. Он быстро насыщается кислородом (намного быстрее, чем гемоглобин), а затем постепенно передает его различным тканям.
Структурные белки выполняют защитную функцию (кожный покров) или опорную скрепляют организм в единое целое и придают ему прочность (хрящи и сухожилия). Их главным компонентом является фибриллярный белок коллаген (рис. 11), наиболее распространенный белок животного мира, в организме млекопитающих, на его долю приходится почти 30% от всей массы белков. Коллаген обладает высокой прочностью на разрыв (известна прочность кожи), но из-за малого содержания поперечных сшивок в коллагене кожи, шкуры животных мало пригодны в сыром виде для изготовления различных изделий. Чтобы уменьшить набухание кожи в воде, усадку при сушке, а также для увеличения прочности в обводненном состоянии и повышения упругости в коллагене создают дополнительные поперечные сшивки (рис. 15а), это, так называемый процесс дубления кожи.
В живых организмах молекулы коллагена, возникшие в процессе роста и развития организма, не обновляются и не замещаются заново синтезированными. По мере старения организма увеличивается количество поперечных сшивок в коллагене, что приводит к снижению его эластичности, а поскольку обновление не происходит, то проявляются возрастные изменения увеличение хрупкости хрящей и сухожилий, появление морщин на коже.
В суставных связках содержится эластин структурный белок, легко растягивающийся в двух измерениях. Наибольшей эластичностью обладает белок резилин, который находится в местах шарнирного прикрепления крыльев у некоторых насекомых.
Роговые образования волосы, ногти, перья состоя, в основном, из белка кератина (рис. 24). Его основное отличие заметное содержание остатков цистеина, образующего дисульфидные мостики, что придает высокую упругость (способность восстанавливать исходную форму после деформации) волосам, а также шерстяным тканям.
Для необратимого изменения формы кератинового объекта нужно вначале разрушить дисульфидные мостики с помощью восстановителя, придать новую форму, а затем вновь создать дисульфидные мостики с помощью окислителя (рис. 16), именно так делается, например, химическая завивка волос.
При увеличении содержания остатков цистеина в кератине и, соответственно, возрастании количества дисульфидных мостиков способность к деформации исчезает, но при этом появляется высокая прочность (в рогах копытных животных и панцирях черепах содержится до 18% цистеиновых фрагментов). В организме млекопитающих содержится до 30 различных типов кератина.
Родственный кератину фибриллярный белок фиброин, выделяемый гусеницами шелкопряда при завивке кокона, а также пауками при плетении паутины, содержит только
b -структуры, соединенные одиночными цепями (рис. 11). В отличие от кератина, у фиброина нет поперечных дисульфидных мостиков, он обладает очень прочен на разрыв (прочность в расчете на единицу поперечного сечения у некоторых образцов паутины выше, чем у стальных тросов). Из-за отсутствия поперечных сшивок фиброин неупруг (известно, что шерстяные ткани почти несминаемы, а шелковые легко мнутся). Регуляторные белки, чаще называемые гормонами, участвуют в различных физиологических процессах. Например, гормон инсулин (рис. 25) состоит из двух a -цепей, соединенных дисульфидными мостиками. Инсулин регулирует обменные процессы с участием глюкозы, его отсутствие ведет к диабету.В гипофизе мозга синтезируется гормон, регулирующий рост организма. Существуют регуляторные белки, контролирующие биосинтез различных ферментов в организме.
Сократительные и двигательные белки придают организму способность сокращаться, изменять форму и перемещаться, прежде всего, речь идет о мышцах. 40% от массы всех белков, содержащихся в мышцах, составляет миозин (mys, myos, греч. мышца). Его молекула содержит одновременно фибриллярную и глобулярную часть (рис. 26)
Такие молекулы объединяются в крупные агрегаты, содержащие 300400 молекул.
При изменении концентрации ионов кальция в пространстве, окружающем мышечные волокна, происходит обратимое изменение конформации молекул изменение формы цепи за счет поворота отдельных фрагментов вокруг валентных связей. Это приводит к сокращению и расслаблению мышц, сигнал для изменения концентрации ионов кальция поступает от нервных окончаний в мышечных волокнах. Искусственное сокращение мышц можно вызвать действием электрических импульсов, приводящих к резкому изменению концентрации ионов кальция, на этом основана стимуляция сердечной мышцы для восстановления работы сердца.
Защитные белки позволяют уберечь организм от вторжения атакующих его бактерий, вирусов и от проникновения чужеродных белков (обобщенное название чужеродных тел антигены). Роль защитных белков выполняют иммуноглобулины (другое их название антитела), они распознают антигены, проникшие в организм, и прочно связываются с ними. В организме млекопитающих, включая человека, есть пять классов иммуноглобулинов: M, G, A, D и E, их структура, как следует из названия, глобулярная, кроме того, все они построены сходным образом. Молекулярная организация антител показана далее на примере иммуноглобулина класса
G (рис. 27). Молекула содержит четыре полипептидные цепи, объединенные тремя дисульфидными мостиками S - S (на рис. 27 они показаны с утолщенными валентными связями и крупными символами S ), кроме того, каждая полимерная цепь содержит внутрицепные дисульфидные перемычки. Две большие полимерные цепи (выделены синим цветом) содержат 400600 аминокислотных остатков. Две другие цепи (выделены зеленым цветом) почти вдвое короче, они содержат приблизительно 220 аминокислотных остатков. Все четыре цепи расположены таким образом, что концевые H 2 N -группы направлены в одну сторону.После контакта организма с чужеродным белком (антигеном), клетки иммунной системы начинают вырабатывать иммуноглобулины (антитела), которые накапливаются в сыворотке крови. На первом этапе основную работу совершают участки цепей, содержащие концевые
H 2 N (на рис. 27 соответствующие участки отмечены светло-синим и светло-зеленым цветом). Это области захвата антигенов. В процессе синтеза иммуноглобулина эти участки формируется таким образом, чтобы их строение и конфигурация максимально соответствовали структуре приблизившегося антигена (как ключ к замку, подобно ферментам, но задачи в данном случае иные). Таким образом, для каждого антигена в качестве иммунного ответа создается строго индивидуальное антитело. Столь «пластично» изменять строение в зависимости от внешних факторов, помимо иммуноглобулинов, не может ни один известный белок. Ферменты решают задачу структурного соответствия реагенту иным путем с помощью гигантского набора разнообразных ферментов в расчете на все возможные случаи, а иммуноглобулины каждый раз заново перестраивают «рабочий инструмент». Сверх того, шарнирный участок иммуноглобулина (рис. 27) обеспечивает двум областям захвата некоторую независимую подвижность, в результате молекула иммуноглобулина может «найти» сразу два наиболее удобных для захвата участка в антигене с тем, чтобы его надежно зафиксировать, это напоминает действия ракообразного существа.Далее включается цепь последовательных реакций иммунной системы организма, подключаются иммуноглобулины других классов, в результате происходит дезактивация чужеродного белка, а затем уничтожение и удаление антигена (постороннего микроорганизма или токсина).
После контакта с антигеном максимальная концентрация иммуноглобулина достигается (в зависимости от природы антигена и индивидуальных особенностей самого организма) в течение нескольких часов (иногда нескольких дней). Организм сохраняет память о таком контакте, и при повторной атаке таким же антигеном иммуноглобулины накапливаются в сыворотке крови значительно быстрее и в большем количестве возникает приобретенный иммунитет.
Приведенная классификация белков носит в определенной степени условный характер, например белок тромбин, упомянутый среди защитных белков, по существу представляет собой фермент , катализирующий гидролиз пептидных связей, то есть, относится к классу протеаз.
К защитным белкам часто относят белки змеиного яда и токсичные белки некоторых растений, поскольку их задача уберечь организм от повреждений.
Есть белки, функции которых настолько уникальны, что это затрудняет их классификацию. Например, белок монеллин, содержащийся в одном из африканских растений, очень сладкий на вкус, он стал предметом изучения как нетоксичное вещество, которое может быть использовано вместо сахара для предотвращения ожирения. Плазма крови некоторых антарктических рыб содержит белки со свойствами антифриза, который предохраняет кровь этих рыб от замерзания.
Искусственный синтез белков. Конденсация аминокислот, приводящая к полипептидной цепи, представляет собой хорошо изученный процесс. Можно провести, например, конденсацию какой либо одной аминокислоты или смеси кислот и получить, соответственно, полимер, содержащий одинаковые звенья, либо различные звенья, чередующиеся в случайном порядке. Такие полимеры мало похожи на природные полипептиды и не обладают биологической активностью. Основная задача состоит в том, чтобы соединять аминокислоты в строго определенном, заранее намеченном порядке, чтобы воспроизвести последовательность аминокислотных остатков в природных белках. Американский ученый Роберт Меррифилд предложил оригинальный метод, позволивший решить такую задачу. Сущность метода состоит в том, что первую аминокислоту присоединяют к нерастворимому полимерному гелю, который содержит реакционно-способные группы, способные соединяться с СООН группами аминокислоты. В качестве такой полимерной подложки был взят сшитый полистирол с введенными в него хлорметильными группами. Чтобы взятая для реакции аминокислота не прореагировала сама с собой и чтобы она не присоединилась H 2 N -группой к подложке, аминогруппу этой кислоты предварительно блокируют объемистым заместителем [(С 4 Н 9) 3 ] 3 ОС(О)-группой. После того, как аминокислота присоединилась к полимерной подложке, блокирующую группу удаляют и в реакционную смесь вводят другую аминокислоту, у которой также предварительно заблокирована H 2 N -группа. В такой системе возможно только взаимодействие H 2 N -группы первой аминокислоты и группы СООН второй кислоты, которое проводят в присутствии катализаторов (солей фосфония). Далее всю схему повторяют, вводя третью аминокислоту (рис. 28).На последней стадии полученные полипептидные цепи отделяют от полистирольной подложки. Сейчас весь процесс автоматизирован, существуют автоматические синтезаторы пептидов, действующие по описанной схеме. Таким методом синтезировано множество пептидов, используемых в медицине и сельском хозяйстве. Удалось также получить улучшенные аналоги природных пептидов с избирательным и усиленным действием. Синтезированы некоторые небольшие белки, например гормон инсулина и некоторые ферменты.
Существуют также методы синтеза белков, копирующие природные процессы: синтезируют фрагменты нуклеиновых кислот, настроенных на получение определенных белков, затем эти фрагменты встраивают в живой организм (например, в бактерию), после чего организм начинает вырабатывать нужный белок. Таким способом сейчас получают значительные количества труднодоступных белков и пептидов, а также их аналогов.
Белки в живом организме постоянно расщепляются на исходные аминокислоты (с непременным участием ферментов), одни аминокислоты переходят в другие, затем белки вновь синтезируются (также с участием ферментов), т.е. организм постоянно обновляется. Некоторые белки (коллаген кожи, волос) не обновляются, организм непрерывно их теряет и взамен синтезирует новые. Белки как источники питания выполняют две основные функции: они поставляют в организм строительный материал для синтеза новых белковых молекул и, кроме того, снабжают организм энергией (источники калорий).Плотоядные млекопитающие (в том числе и человек) получают необходимые белки с растительной и животной пищей. Ни один из полученных с пищей белков не встраивается в организм в неизменном виде. В пищеварительном тракте все поглощенные белки расщепляются до аминокислот, и уже из них строятся белки, необходимые конкретному организму, при этом из 8 незаменимых кислот (табл. 1) в организме могут синтезироваться остальные 12, если они не поступают в достаточном количестве с пищей, но незаменимые кислоты должны поступать с пищей непременно. Атомы серы в цистеине организм получает с незаменимой аминокислотой метионином. Часть белков распадается, выделяя энергию, необходимую для поддержания жизнедеятельности, а содержавшийся в них азот выводится из организма с мочой. Обычно организм человека теряет 2530 г. белка в сутки, поэтому белковая пища должны постоянно присутствовать в нужном количестве. Минимальная суточная потребность в белке составляет у мужчин 37 г, у женщин 29 г, однако рекомендованные нормы потребления почти вдвое выше. При оценке пищевых продуктов важно учитывать качество белка. При отсутствии или низком содержании незаменимых аминокислот белок считается малоценным, поэтому такие белки должны потребляться в большем количестве. Так, белки бобовых культур содержат мало метионина, а в белках пшеницы и кукурузы низкое содержанием лизина (обе аминокислоты незаменимые). Животные белки (исключая коллагены) относят к полноценным пищевым продуктам. Полный набор всех незаменимых кислот содержит казеин молока, а также приготовляемые из него творог и сыр, поэтому вегетарианская диета, в том случае, если она очень строгая, т.е. «безмолочная», требует усиленного потребления бобовых культур, орехов и грибов для снабжения организма незаменимыми аминокислотами в нужном количестве.
Синтетические аминокислоты и белки используют и как пищевые продукты, добавляя их в корма, которые содержат незаменимые аминокислоты в малом количестве. Существуют бактерии, которые могут перерабатывать и усваивать углеводороды нефти, в этом случае для полноценного синтеза белков их нужно подкармливать азотсодержащими соединениями (аммиак или нитраты). Получаемый таким способом белок используют в качестве корма для скота и домашней птицы. В комбикорм домашних животных часто добавляют набор ферментов карбогидраз, которые катализируют гидролиз трудно разлагаемых компонентов углеводной пищи (клеточные стенки зерновых культур), в результате чего растительная пища усваивается более полно.
Михаил Левицкий
ЛИТЕРАТУРА Шульц Г., Ширмер Р. Принципы структурной организации белков . М., Мир, 1982Овчинников Ю.А. Биоорганическая химия , М., Просвещение, 1987
Борисов В.В. Каждый белок свой сюжет . Химия и жизнь. 1990, № 2
Иванов В.И. Как работают ферменты . Соросовский образовательный журнал. 1996, № 9
