Жаберное дыхание для земноводных характерно. Интересные факты о земноводных
Обмен газами, или дыхание, выражается в поглощении организмом кислорода из окружающей среды (воды или атмосферы) и выделении в последнюю углекислого газа как конечного продукта протекающего в тканях окислительного процесса, благодаря которому освобождается необходимая для жизнедеятельности энергия. Кислород воспринимается организмом различными способами; их в основном можно характеризовать как: 1) диффузное дыхание и 2) дыхание местное, т. е. специальными органами.
Диффузное дыхание заключается в поглощении кислорода и выделении углекислого газа всей поверхностью наружного покрова-к о ж н о е д ы-х а н и е-и эпителиальной оболочкой пищеварительной трубки-к и ш е ч-н о е дыхание, т. е. без специально приспособленных для этого органов. Подобный способ газообмена свойствен некоторым типам примитивных многоклеточных животных, как, например, губкам, кишечнополостным и плоским червям, и обусловливается отсутствием у них системы кровообращения.
Само собой понятно, что диффузное дыхание присуще только организмам, у которых объём тела мал, а поверхность его относительно обширна, так как известно, что объём тела возрастает пропорционально кубу радиуса, а соответствующая поверхность - только квадрату радиуса. Следовательно, при большом объёме тела такой способ дыхания оказывается недостаточным.
Однако и при более или менее соответствующих отношениях объёма к поверхности диффузное дыхание всё же не всегда может удовлетворять организмы, так как чем энергичнее проявляется жизнедеятельность, тем интенсивнее должны протекать окислительные процессы в теле.
При интенсивных проявлениях жизни, несмотря на малый объём тела, необходимо увеличение его площади соприкосновения со средой, содержащей кислород, и особые приспособления для ускорения вентилирования дыхательных путей. Увеличение площади обмена газами достигается развитием специальных органов дыхания.
Специальные органы дыхания значительно варьируют в деталях построения и местоположения в организме. Для водных животных такими органами служат жабры, для наземных-т р а х е и у беспозвоночных, а у позвоночных- лёгкие.
Жаберное дыхание. Жабры бывают наружные и внутренние. Примитивные наружные жабры представляют простое выпячивание ворсинчатых отпрысков кожного покрова, обильно снабжённых капиллярными сосудами. Такие жабры в некоторых случаях своей функцией мало чем отличаются от диффузного дыхания, являясь лишь более высокой его ступенью (рис. 332-А, 2). Обычно они концентрируются в передних участках тела.
Внутренние жабры формируются из складок слизистой оболочки начального участка пищеварительной трубки между жаберными щелями (рис. 246-2-5; 332-7). Прилежащий к ним кожный покров образует обильные разветвления в виде лепестков с большим количеством капиллярных кровеносных сосудов. Внутренние жабры часто бывают прикрыты специальной складкой кожного покрова (жаберной покрышкой), колебательные движения которой улучшают условия обмена, усиливая приток воды и удаляя использованные её порции.
Внутренние жабры свойственны водным позвоночным, причём акт газообмена у них усложняется пропуском порций воды к жаберным щелям через ротовую полость и движениями жаберной покрышки. Кроме того, жабры у них включены в круг кровообращения. К каждой жаберной дуге подходят свои сосуды, и, таким образом, одновременно осуществляется более высокая ди-ференциация системы кровообращения.
Разумеется, при жаберных способах газообмена может сохраняться и кожное дыхание, но настолько слабое, что оно отодвигается на задний план.
При описании ротоглотки пищеварительного тракта уже было сказано, что жаберный аппарат свойствен и некоторым беспозвоночным, как, например, полухордовым и хордовым животным.
Лёгочное дыхание -весьма совершенный способ газообмена, легко обслуживающий организмы массивных животных. Оно свойственно наземным позвоночным: амфибиям (не в личиночном состоянии), рептилиям, птицам и млекопитающим. К акту газообмена, сосредоточенного в лёгких, присоединяется ряд органов с иными функциями, вследствие чего лёгочный способ дыхания требует развития очень сложного комплекса органов.
Сравнивая водные и наземные типы дыхания позвоночных, следует иметь в виду одно важное анатомическое их различие. При жаберном дыхании порции воды одна за другой поступают в примитивный рот и выпускаются через жаберные щели, где сосудами жаберных складок из неё извлекается кислород. Таким образом, аппарату жаберного дыхания позвоночных свойственны входное и ряд выходных отверстий. При лёгочном дыхании для введения и выведения воздуха используются одни и те же отверстия. Эта особенность, естественно, связана с необходимостью вбирать и выталкивать порции воздуха для более быстрого вентилирования площади газообмена, т. е. с необходимостью расширения и сужения лёгких.
Можно допустить, что у отдалённых, более примитивных предков позвоночных в стенках преобразующегося в лёгкое плавательного пузыря была самостоятельная мускульная ткань; её периодическими сокращениями воздух выталкивался из пузыря, а в результате его расправления в силу эластичности пузырных стенок набирались свежие порции воздуха. Эластическая ткань, наряду с хрящевой, и теперь господствует в качестве опоры в органах дыхания.
В дальнейшем, с повышением жизнедеятельности организмов, такой механизм дыхательных движений становился уже несовершенным. В истории развития он был замещён силой, сосредоточенной или в ротовой полости и переднем участке трахеи (амфибии), или в стенках грудной и брюшной полостей (рептилии, млекопитающие) в виде специально диференцирован-ной части туловищной мускулатуры (дыхательные мускулы) и, наконец, диафрагмы. Лёгкое подчиняется движениям этой мускулатуры, расширяясь и суживаясь пассивно, и сохраняет необходимую для этого эластичность, а также небольшой мышечный аппарат как подсобное приспособление.
 Кожное дыхание становится настолько ничтожным, что роль его сводится почти к нулю.
Кожное дыхание становится настолько ничтожным, что роль его сводится почти к нулю.
Газообмен в лёгких у наземных позвоночных, так же как у водных, теснейшим образом связан с системой кровообращения посредством организации обособленного, дыхательного, или малого, круга кровообращения.
Вполне понятно, что основные структурные изменения в организме при лёгочном дыхании сводятся: 1) к увеличению соприкосновения рабочей площади лёгких с воздухом и 2) к весьма тесной и не менее обширной связи этой площади с тонкостенными капиллярами круга кровообращения.
Функция дыхательного аппарата-пропускать в свои многочисленные каналы воздух для газообмена-говорит за характер его построения в виде открытой, зияющей системы трубок. Их стенки, по сравнению с мягкой кишечной трубкой, слагаются из более твёрдого опорного материала; местами в виде костной ткани (носовая полость), а главным образом в виде хрящевой ткани и легко податливой, но быстро возвращающейся к норме упругой ткани.
Слизистая оболочка дыхательных путей выстлана специальным мерцательным эпителием. Лишь на немногих участках он видоизменяется в другую форму в соответствии с другими функциями данных участков, как, например, в обонятельной области и в местах самого газообмена.
На протяжении лёгочного дыхательного тракта останавливают на себе внимание три своеобразных участка. Из них начальный-н осовая поло с т ь-служит для воспринятая воздуха, обследуемого здесь на запах. Второй участок-г о р т а н ь-является приспособлением для изолирования дыхательного тракта от пищеварительного пути при проходе пищевого кома через глотку, для издавания звуков и, наконец, для производства кашлевых толчков, выбрасывающих из дыхательных путей слизь. Последний участок-л ё г к и е-представляют орган непосредственного обмена газами.
Между носовой полостью и гортанью располагается общая с пищеварительным аппаратом полость глотки, а между гортанью и лёгким тянется дыха-
тельное горло, или трахея. Таким образом, проходящий воздух используется описанными расширяющимися участками в трёх различных направлениях: а) воспринятая запахов, б) приспособления для издавания звуков и, наконец, в) газообмена, из которых последнее-основное.
Класс Земноводные = Амфибии.
Первые наземные позвоночные, еще сохранившие связь с водной средой. Класс насчитывает 3900 видов и включает 3 отряда: хвостатые (саламандры, тритоны), безногие (тропические червяги) и бесхвостые (жабы, квакши, лягушки, и др.).
Вторично-водные животные. Поскольку в яйце отсутствует амниотическая полость (вместе с круглоротыми и рыбами амфибии относятся к анамниям), они размножаются в воде, где проходят начальные этапы их развития. На разных стадиях жизненного цикла земноводные ведут наземный или полуводный образ жизни, распространены практически повсеместно, преимущественно в районах с высокой влажностью по берегам пресных водоемов и на сырых почвах. Среди земноводных нет форм, которые могли бы жить в соленой морской воде. Характерны различные способы передвижения: известны виды, совершающие довольно длинные прыжки, передвигающиеся шагом или «ползающие», лишенные конечностей (червяги).
Основные признаки земноводных.
Амфибии сохранили многие черты своих чисто водных предков, но наряду с этим приобрели и ряд признаков, свойственных настоящим наземным позвоночным.
Для хвостатых и бесхвостых характерно личиночное развитие с жаберным дыханием в пресной воде (головастики лягушки) и их метаморфоз во взрослую особь, дышащую легкими. У безногих по вылуплении личинка принимает вид взрослого животного.
Кровеносная система характеризуется двумя кругами кровообращения. Сердце трехкамерное. Имеет один желудочек и два предсердия.
Обособляются шейный и крестцовый отделы позвоночника, имеющие по одному позвонку.
Для взрослых земноводных характерны парные конечности с шарнирными суставами. Конечности пятипалые.
Череп двумя затылочными мыщелками подвижно сочленяется с шейным позвонком.
Тазовый пояс плотно причленяется к поперечным отросткам крестцового позвонка.
Глаза имеют подвижные веки и мигательные перепонки для предохранения глаз от засорения и высыхания. За счет выпуклой роговицы и уплощенного хрусталика улучшается аккомодация.
Передний мозг увеличивается и разделяется на два полушария. Средний мозг и мозжечок развиты незначительно. От головного мозга отходит 10 пар черепно – мозговых нервов.
Кожа голая, т.е. лишена каких – либо роговых или костных образований, проницаема для воды и газов. Поэтому она всегда влажная – кислород сначала растворяется в жидкости, покрывающей кожу, после чего диффундирует в кровь. То же самое происходит с углекислым газом, но в обратном направлении.
Почки, как у рыб, первичные = мезонефрические.
Для улавливания звуковых волн воздушной среды появляется барабанная перепонка, за ней - среднее ухо (барабанная полость), в котором располагается слуховая косточка – стремя, которая проводит колебания к внутреннему уху. Евстахиевой трубой полость среднего уха сообщается с ротовой полостью. Появляются хоаны – внутренние ноздри, носовые ходы становятся сквозными.
Температура тела непостоянная (пойкилотермия) зависит от температуры окружающей среды и лишь слегка превышает последнюю.
Ароморфозы:
Появились легкие и легочное дыхание.
Усложнилась кровеносная система, развился легочный круг кровообращения, т.е. у амфибий два круга кровообращения – большой и малый. Сердце трехкамерное.
Сформировались парные пятипалые конечности, представляющие собой систему рычагов с шарнирными суставами и предназначенные для передвижения по суше.
Образовался шейный отдел в позвоночнике, обеспечивающий движение головы, и крестцовый отдел – место прикрепления тазового пояса.
Появились среднее ухо, веки, хоаны.
Дифференцировка мышц.
Прогрессивное развитие нервной системы.
Филогения.
Земноводные произошли от древних кистеперых рыб в девонском периоде палеозойской эры примерно 350 млн. лет назад. Первые земноводные – ихтиостеги – по внешнему виду напоминали современных хвостатых амфибий. В их строении были черты, свойственные рыбам, в том числе рудименты жаберной крышки и органы боковой линии.
Покров. Двухслойный. Эпидермис многослойный, кориум – тонкий, но обильно снабжен капиллярами. У земноводных сохранилась способность вырабатывать слизь, но не отдельными клетками, как у большинства рыб, а оформленными слизистыми железами альвеолярного типа. Кроме того, у амфибий часто имеются зернистые железы с ядовитым секретом разной степени токсичности. Цвет кожи амфибий зависит от особых клеток – хроматофоров. К их числу принадлежат меланофоры, липофоры и иридоциты.
Под кожей у лягушек имеются обширные лимфатические лакуны – резервуары, заполненные тканевой жидкостью и позволяющие при неблагоприятных условиях накапливать запас воды.
Скелет подразделяется на осевой и добавочный, как у всех позвоночных. Позвоночный столб более дифференцирован на отделы, чем у рыб и состоит из четырех отделов: шейного, туловищного, крестцового и хвостового. Шейный и крестцовый отделы имеют по одному позвонку. Туловищных позвонков у бесхвостых обычно семь, а все хвостовые позвонки (примерно 12) сливаются в единую косточку – уростиль. У хвостатых 13 – 62 туловищных и 22 – 36 хвостовых позвонков; у безногих общее число позвонков доходит до 200 – 300. Наличие шейного позвонка важно, т.к. в отличие от рыб, земноводные не могут так быстро разворачивать свое тело, а шейный позвонок делает голову подвижной, но с малой амплитудой. Амфибии не могут поворачивать голову, но могут ее наклонять.
Позвонки у разных амфибий могут различаться по своему типу. У безногих и низших хвостатых позвонки амфицельные, с сохранившейся хордой, как у рыб. У высших хвостатых позвонки опистоцельные, т.е. спереди тела выгнуты, а сзади вогнуты. У бесхвостых, наоборот, передняя поверхность тел позвонков вогнута, а задняя выгнута. Такие позвонки называются процельными. Наличие сочленовных поверхностей и сочленовных отростков обеспечивает не только прочное соединение позвонков, но и делает осевой скелет подвижным, что важно для передвижения хвостатых амфибий в воде без участия конечностей, за счет боковых изгибов тела. Кроме того, возможны вертикальные движения.
Череп амфибий представляет собой как бы видоизмененный череп костистой рыбы, приспособленный к наземному существованию. Мозговой череп пожизненно сохраняется преимущественно хрящевым. Затылочная область черепа содержит только две боковые затылочные кости, которые несут по сочленовному мыщелку, с помощью которых череп крепится к позвонкам. Висцеральный череп амфибий претерпевает наибольшие преобразования: возникают вторичные верхние челюсти; образованные межчелюстными (предчелюстными) и верхнечелюстными костями. Редукция жаберного дыхания привела к радикальному изменению подъязычной дуги. Подъязычная дуга преобразуется в элемент слухового аппарата и подъязычную пластинку. В отличие от рыб, висцеральный череп амфибий непосредственно прирастает небно-квадратным хрящом ко дну мозгового черепа. Такой тип прямого соединения компонентов черепа без участия элементов подъязычной дуги называется аутостилией. Элементы жаберной крышки у амфибий отсутствуют.
Добавочный скелет включает в себя кости поясов и свободных конечностей. Как и у рыб, кости плечевого пояса амфибий находятся в толще мышц, которые связывают их с осевым скелетом, но сам пояс не связан с осевым скелетом непосредственно. Пояс обеспечивает опору для свободной конечности.
Всем наземным животным постоянно приходится преодолевать силу тяжести, чего не нужно делать рыбам. Свободная конечность служит опорой, позволяет приподнять тело над поверхностью и обеспечивает перемещение. Свободные конечности состоят из трех отделов: проксимального (одна кость), промежуточного (две кости) и дистального (относительно большое число костей). У представителей разных классов наземных позвоночных встречаются особенности строения той или иной свободной конечности, но все они носят вторичный характер.
У всех амфибий проксимальный отдел свободной передней конечности представлен плечевой костью, промежуточный – локтевой и лучевой костями у хвостатых и единой костью предплечья (она образована в результате сращения локтевой и лучевой кости) у бесхвостых. Дистальный отдел образован запястью, пястью и фалангами пальцев.
Пояс задних конечностей сочленяется непосредственно с осевым скелетом, с его крестцовым отделом. Надежное и жесткое соединение тазового пояса с позвоночным столбом обеспечивает работу задних конечностей, которые более важны для перемещения амфибий.
Мышечная система отличается от мышечной системы рыб. Туловищная мускулатура сохраняет метамерное строение только у безногих. У хвостатых метамерия сегментов нарушается, а у бесхвостых амфибий начинают обособляться участки мышечных сегментов, дифференцируясь в лентовидные мышцы. Резко возрастает масса мускулатуры конечностей. У рыб движения плавников обеспечиваются главным образом мышцами, расположенными на туловище, тогда как пятипалая конечность двигается за счет мышц, расположенной в ней самой. Появляется сложная система мускулов – антагонистов – мышц сгибателей и разгибателей. Сегментированная мускулатура имеется только в области позвоночного столба. Усложняется и специализируется мускулатура ротовой полости (жевательная, языка, дна ротовой полости), не только участвующая в захвате и проглатывании пищи, но и обеспечивающая вентиляцию ротовой полости и легких.
Полость тела – целом. У земноводных в связи с исчезновением жабр изменилось относительное положение перикардиальной полости. Ее вытолкнуло на дно грудной клетки в зону, прикрывающуюся грудиной (или коракоидом). Над ней в паре целомических каналов лежат легкие. Полости, вмещающие сердце и легкие. Разделяет плеврокардиальная перепонка. Полость, в которой располагаются легкие, сообщается с главным целомом.
Нервная система. Головной мозг ихтиопсидного типа, т.е. главным интегрирующим центром является средний мозг, но мозг амфибий имеет ряд прогрессивных изменений. Мозг амфибий имеет пять отделов и отличается от мозга рыб, главным образом, большим развитием переднего мозга, полным разделением его полушарий. Кроме того, нервное вещество уже выстилает кроме дна боковых желудочков также бока и крышу, образуя мозговой свод – архипаллиум. Развитие архипаллиума, сопровождающееся усилением связей с промежуточным и особенно средним мозгом, приводит к тому, что ассоциативная деятельность, регулирующая поведение, осуществляется у земноводных не только продолговатым и средним мозгом, но и полушариями переднего мозга. Удлиненные полушария спереди имеют общую обонятельную долю, от которой берут начало два обонятельных нерва. Позади переднего мозга находится промежуточный мозг. На его крыше располагается эпифиз. На нижней стороне мозга имеется перекрест зрительных нервов (хиазма). От дна промежуточного мозга отходит воронка и гипофиз (нижняя мозговая железа).
Средний мозг представлен в виде двух круглых зрительных долей. Сзади зрительных долей лежит слаборазвитый мозжечок. Сразу же за ним расположен продолговатый мозг с ромбовидной ямкой (четвертый желудочек). Продолговатый мозг постепенно переходит в спинной мозг.
У земноводных от головного мозга отходит 10 пар головных нервов. Одиннадцатая пара не развита, а двенадцатая отходит за пределами черепа.
Настоящих спинномозговых нервов у лягушки 10 пар. Три передние принимают участие в образовании плечевого сплетения, иннервирующего передние конечности, а четыре задние пары – в образовании пояснично-крестцового сплетения, иннервирующего задние конечности.
Органы чувств обеспечивают ориентировку земноводных в воде и на суше.
Органы боковой линии есть у всех личинок и у взрослых с водным образом жизни. Представлены скоплением чувствительных клеток с подходящими к ним нервами, которые разбросаны по всему телу. Чувствительные клетки воспринимают температурные, болевые, тактильные ощущения, а также изменение влажности и химического состава окружающей среды.
Органы обоняния. У амфибий с каждой стороны головы имеется небольшая наружная ноздря, которая ведет в удлиненный мешок, заканчивающийся внутренней ноздрей (хоаной). Хоаны открываются в передней части крыши ротовой полости. Спереди от хоан слева и справа имеется по мешку, которые открываются в носовую полость. Это т.н. вомероназальный орган. В нем большое количество сенсорных клеток. Его функция состоит в получении обонятельной информации о пище.
Органы зрения имеют строение, характерное для наземного позвоночного. Это выражается в выпуклой форме роговицы, хрусталике в виде двояковыпуклой линзы, в подвижных веках, защищающих глаза от высыхания. Но аккомодация, как и у рыб, достигается перемещением хрусталика путем сокращения ресничной мышцы. Мышца располагается в кольцевом валике, окружающем хрусталик, и при ее сокращение хрусталик лягушки несколько выдвигается вперед.
Орган слуха устроен по наземному типу. Появляется второй отдел – среднее ухо, в котором помещается впервые появляющаяся у позвоночных слуховая косточка – стремя. Барабанная полость соединена с глоточной областью евстахиевой трубой.
Поведение земноводных весьма примитивно, условные рефлексы вырабатываются медленно, а угасают быстро. Двигательная специализация рефлексов очень мала, поэтому лягушка не может сформировать защитный рефлекс отдергивания одной лапки, и при раздражении одной конечности она дергает обеими лапками.
Пищеварительная система начинается ротовой щелью, ведущей в ротоглоточную полость. В ней располагается мускулистый язык. В нее открываются протоки слюнных желез. Язык и слюнные железы впервые появляются у амфибий. Железы служат только для смачивания пищевого комка и в химической обработке пищи не участвуют. На межчелюстных, верхнечелюстных костях, сошнике располагаются простые конические зубы, которые основанием прикрепляются к кости. Пищеварительная трубка дифференцируется на ротоглоточную полость, короткий пищевод, выполняющий функцию проведения пищи в желудок, и объемистый желудок. Пилорическая его часть переходит в двенадцатиперстную кишку – начало тонкого кишечника. В петле между желудком и двенадцатиперстной кишкой лежит поджелудочная железа. Тонкая кишка плавно переходит в толстую кишку, которая заканчивается выраженной прямой кишкой, открывающейся в клоаку.
Пищеварительными железами являются печень с желчным пузырем и поджелудочная железа. Протоки печени вместе с протоком желчного пузыря открываются в двенадцатиперстную кишку. Протоки поджелудочной железы впадают в проток желчного пузыря, т.е. самостоятельного сообщения с кишечником эта железа не имеет.
Т.о. пищеварительная система амфибий отличается от аналогичной системы рыб большей длиной пищеварительного тракта, конечный отдел толстой кишки открывается в клоаку.
Кровеносная система замкнутая. Два круга кровообращения. Сердце трехкамерное. Кроме того, в сердце имеется венозный синус, сообщающийся с правым предсердием, а с правой стороны желудочка отходит артериальный конус. От него отходят три пары сосудов, гомологичных жаберным артериям рыб. Каждый сосуд начинается самостоятельным отверстием. Все три сосуда левой и правой стороны идут вначале общим артериальным стволом, окруженным общей оболочкой, а затем разветвляются.
Сосуды первой пары (считая от головы), гомологичные сосудам первой пары жаберных артерий рыб, называются сонными артериями, которые несут кровь к голове. По сосудам второй пары (гомологичным второй паре жаберных артерий рыб) – дугам аорты – кровь направляется к задней части тела. От дуг аорты отходят подключичные артерии, несущие кровь к передним конечностям.
По сосудам третьей пары, гомологичным четвертой паре жаберных артерий рыб – легочным артериям – кровь направляется в легкие. От каждой легочной артерии отходит крупная кожная артерия, по которой кровь направляется в кожу для окисления.
Венозная кровь от переднего конца тела собирается по двум парам яремных вен. Последняя, сливаясь с кожными венами, уже принявшими в себя подключичные вены, образует две передние полые вены. Они несут в венозный синус смешанную кровь, поскольку по кожным венам движется артериальная кровь.
У личинок земноводных один круг кровообращения, их кровеносная система подобна кровеносной системе рыб.
У амфибий появляется новый орган кровообращения – красный костный мозг трубчатых костей. Эритроциты крупные, ядерные, лейкоциты неодинаковы по внешнему виду. Есть лимфоциты.
Лимфатическая система. Кроме лимфатических мешков, расположенных под кожей, имеются лимфатические сосуды и сердца. Одна пара лимфатических сердец помещается около третьего позвонка, другая – вблизи клоачного отверстия. Селезенка, имеющая вид небольшого круглого тела красного цвета, располагается на брюшине вблизи начала прямой кишки.
Дыхательная система. Принципиально отличается от дыхательной системы рыб. У взрослых особей органами дыхания являются легкие и кожа. Дыхательные пути в связи с отсутствием шейного отдела короткие. Представлены носовой и ротоглоточной полостями, а также гортанью. Гортань открывается непосредственно в легкие двумя отверстиями. Вследствие редукции ребер легкие наполняются посредством заглатывания воздуха – по принципу нагнетательного насоса.
Анатомически дыхательная система амфибий включает в себя ротоглоточную полость (верхние воздухоносные пути) и гортанно – трахеальную полость (нижние пути), которая непосредственно переходит в мешковидные легкие. Легкое в процессе эмбрионального развития образуются как слепой вырост переднего (глоточного) отдела пищеварительной трубки, поэтому и во взрослом состоянии остается связанным с глоткой.
Т.о. дыхательная система у наземных позвоночных анатомически и функционально подразделяется на два отдела – систему воздухоносных путей и респираторный отдел. Воздухоносные пути осуществляют двусторонний транспорт воздуха, но не участвуют в самом газообмене, респираторный отдел осуществляет газообмен между внутренней средой организма (кровью) и атмосферным воздухом. Газообмен происходит через поверхностную жидкость и идет пассивно в соответствии с градиентом концентрации.
Система жаберных крышек становится ненужной, поэтому жаберный аппарат у всех наземных животных частично видоизменяется, его скелетные структуры частично входят в состав скелета (хрящей) гортани. Вентиляция легких осуществляется за счет принудительных движений специальной соматической мускулатуры в процессе дыхательного акта.
Выделительная система, как и у рыб, представлена первичными, или туловищными почками. Это компактные тела красновато – коричневого цвета, лежащие по бокам позвоночника, а не лентовидные, как у рыб. От каждой почки тянется к клоаке тонкий вольфов канал. У самок лягушек он служит только мочеточником, а у самцов и мочеточником и семяпроводом. В клоаку вольфовы каналы открываются самостоятельными отверстиями. Также отдельно открывается в клоаку и мочевой пузырь. Окончательным продуктом азотистого обмена у амфибий является мочевина. У водных личинок амфибий основным продуктом азотистого обмена является аммиак, который в виде раствора выводится через жабры и кожу.
Амфибии – гиперосмотические животные по отношению к пресной воде. Вследствие этого вода постоянно поступает в организм через кожу, которая не имеет механизмов, препятствующих этому, как у других наземных позвоночных. Морская вода гиперосмотична по отношению к осмотическому давлению в тканях земноводных, при помещении их в такую среду вода через кожу будет уходить из организма. Вот почему земноводные не могут жить в морской воде и погибают в ней от обезвоживания.
Половая система. У самцов органы размножения представлены парой округлых беловатых семенников, прилегающих к брюшной поверхности почек. От семенников к почкам тянутся тонкие семявыводящие канальцы. Половые продукты из семенника через эти канальцы направляются в тела почек, далее в вольфовы каналы и по ним в клоаку. Перед впадением в клоаку вольфовы каналы образуют небольшое расширение – семенные пузырьки, служащие для временного депонирования спермы.
Органы размножения самок представлены парными яичниками зернистого строения. Сверху над ними располагаются жировые тела. В них накапливаются питательные вещества, обеспечивающие формирование половых продуктов во время зимней спячки. В боковых частях полости тела расположены сильно извитые светлые яйцеводы, или мюллеровы каналы. Каждый яйцевод в полость тела в области сердца открывается воронкой; нижняя маточная часть яйцеводов резко расширена и открывается в клоаку. Созревшие яйца через разрыв стенок яичника выпадают в полость тела, затем захватываются воронками яйцеводов и по ним перемещаются в клоаку.
Вольфовы каналы у самок выполняют только функции мочеточников.
У бесхвостых земноводных оплодотворение наружное. Икринка сразу же орошаются семенной жидкостью.
Наружные половые признаки самцов:
У самцов на внутреннем пальце передних конечностей имеется половая бородавка, которая достигает особенного развития ко времени размножения и помогает самцам удерживать самок при оплодотворении икры.
Самцы обычно меньше самок.
Развитие земноводных сопровождается метаморфозом. Яйца содержат относительно немного желтка (мезолецитальные яйца), поэтому происходит радиальное дробление. Из яйца выходит личинка – головастик, которая по своей организации значительно ближе к рыбам, чем к взрослым амфибиям. Он имеет характерную рыбообразную форму - длинный хвост, окруженный хорошо развитой плавательной перепонкой, по бокам головы у него есть две – три пары наружных перистых жабр, парные конечности отсутствуют; имеются органы боковой линии, функционирующей почкой является пронефрос (предпочка). Вскоре наружные жабры исчезают, а взамен их развиваются три пары жаберных щелей с их жаберными лепесточками. В это время сходством головастика с рыбой является также двухкамерное сердце, один круг кровообращения. Затем путем выпячивания из брюшной стенки пищевода развиваются парные легкие. На этой стадии развития артериальная система головастика чрезвычайно похожа на артериальную систему кистеперых и двоякодышащих рыб, и все отличие сводится к тому, что благодаря отсутствию четвертой жабры четвертая приносящая жаберная артерия без перерыва переходит в легочную артерию. Еще позже жабры редуцируются. Впереди жаберных щелей с каждой стороны образуется складка кожи, которая, постепенно разрастаясь назад, затягивает эти щели. Головастик переходи всецело на легочное дыхание и заглатывает ртом воздух. В дальнейшем у головастика формируется парные конечности – сперва передние, потом задние. Однако передние находятся дольше скрытыми под кожей. Хвост и кишечник начинают укорачиваться, появляется мезонефрос, личинка постепенно переходит от растительной пищи к животной и превращается в молодую лягушку.
В ходе развития личинки у нее перестраиваются внутренние системы: дыхательная, кровеносная, выделительная, пищеварительная. Метаморфоз завершается формированием миниатюрной копии взрослой особи.
Для амбистом характерная неотения, т.е. у них размножаются личинки, которых длительное время принимали за самостоятельный вид, поэтому у них есть свое название – аксолотль. Такая личинка крупнее взрослой особи. Другой интересной группой являются протеи, постоянно живущие в воде, которые в течение всей жизни сохраняют наружные жабры, т.е. признаки личинки.
Метаморфоз головастика в лягушку представляет огромный теоретический интерес, т.к. не только доказывает, что земноводные произошли от рыбообразных существ, но дает возможность в подробностях восстановить эволюцию отдельных систем органов, в частности систем кровообращения и дыхания, при переходе водных животных в наземные.
Значение земноводных состоит в том, что они поедают многих вредных беспозвоночных и сами служат пищей другим организмам в цепях питания.
Совокупность процессов, обеспечивающих в организме потребление O 2 и выделение CO 2 , называется дыханием . Различают процессы внешнего и внутреннего дыхания. Внешнее дыхание обеспечивает обмен газов между организмом и внешней средой, внутреннее дыхание - потребление O2 и выделение CO 2 клетками организма.
Фактором, обеспечивающим диффузию газов через дыхательные поверхности, является разность их концентраций. Движение растворенных газов происходит в направлении из области с их высокой концентрацией в область низкой концентрации.
У мелких организмов газообмен, как правило, осуществляется диффузно всей поверхностью тела (или клетки). У более крупных животных газы транспортируются к тканям либо непосредственно (трахейная система насекомых), либо с помощью специальных транспортных средств (кровь, гемолимфа).
Количество кислорода, поступающее в ткани животного, зависит от площади дыхательной поверхности и разности концентрации кислорода на них. Поэтому во всех органах дыхании наблюдается разрастание дыхательного эпителия. Для поддержания же высокого градиента диффузии кислорода на обменной мембране необходимо движение среды (вентиляции). Оно обеспечивается дыхательными ритмическими движениями всего тела животного (малощетинковый червь трубочник, пиявки) либо определенных его участков (ракообразные), а также работой ресничного эпителия (моллюски, ланцетник).
Ряд достаточно крупных животных не имеет специализированных органов дыхания. У них газообмен осуществляется через влажные кожные покровы, снабженные обильной сетью кровеносных сосудов (дождевой червь). Кожное дыхание в качестве дополнительного характерно для животных, обладающих специализированными органами дыхания. Например, у угрей, имеющих жабры, потребность в кислороде на 60% обеспечивается за счет кожного дыхания, у лягушек, имеющих легкие, эта величина составляет более 50 %.
Органами дыхания в водной среде являются жабры, в наземно-воздушной - легкие и трахеи.
Жабры представляют собой органы, расположенные вне полости тела в виде эпителиальных поверхностей, пронизанных густой сетью кровеносных капилляров. Жаберное дыхание свойственно многощетинковым кольчатым червям, большинству моллюсков, ракообразным, рыбам, личинкам земноводных. Наиболее эффективно жаберное дыхание у рыб. В его основе лежит явление противотока : кровь в капиллярах жаберных лепестков течет в направлении, противоположном току волы, омывающей жабры.
Легкие , как правило, являются внутренними органами и защищены от высыхания. Различают два их типа: диффузионные и вентиляционные . В легких первого типа газообмен осуществляется только путем диффузии. Такие легкие имеют относительно небольшие животные: легочные моллюски, скорпионы, пауки. Вентиляционные легкие имеют только наземные позвоночные.
Усложнение строения легких в ряду от земноводных к млекопитающим связано с возрастанием площади дыхательного эпителия. Так, у земноводных 1 см 3 легочной ткани имеет общую газообменную поверхность 20 см 2 . Аналогичный показатель для эпителия легких человека равен 300 см 2 .
Одновременно с увеличением дыхательной поверхности происходит совершенствование механизма вентиляции легких, которая, начиная с пресмыкающихся, осуществляется за счет изменения объема грудной клетки, а у млекопитающих - с участием мышц диафрагмы. Эти приспособления позволили теплокровным (птицам и млекопитающим) резко повысить интенсивность метаболизма.
Третий тип органов дыхания - трахеи . Они представляют собой заполненные воздухом тонкостенные, ветвящиеся неспадающиеся впячивания внутрь тела. Трахеи сообщаются с наружной средой отверстиями в кутикуле - дыхальцами. У насекомых их чаще всего 12 пар: 3 пары на груди и 9 пар на брюшке. Дыхальца могут закрываться либо открываться в зависимости от количества кислорода. При высокой степени развития трахейной системы (у насекомых) ее многочисленные разветвления оплетают все внутренние органы и непосредственно обеспечивают газообмен в тканях. Принципиальное отличие трахейного дыхания от легочного и жаберного заключается в том, что оно не нуждается в участии крови как транспортного посредника в газообмене.
Трахейная система способна поддерживать достаточно высокий уровень тканевого дыхания, обеспечивая тем самым высокую физиологическую активность насекомого.
Вентиляция трахей у насекомых в отсутствие полета осуществляется чаще всего ритмическими сокращениями брюшка, в при полете усиливается движениями груди.
Водные личинки некоторых насекомых дышат при помощи трахейных жабер . В этом случае трахейная система лишена дыхалец т.е. она замкнута и заполнена воздухом. Ветви замкнутой трахейной системы заходят в «жабры» - придатки с большой поверхностью и тонкой кутикулой, позволяющей осуществлять газообмен между водой и воздухом трахейной системы. Такие трахейные жабры есть, например, у личинок поденок. У личинок некоторых стрекоз трахейные жабры расположены в полости прямой кишки, и насекомое вентилирует их, набирая воду внутрь кишки и выталкивая ее обратно.
У амфибий имеются два рода дыхательных органов (не считая кожи): жабры и легкие. Ослабление жаберного дыхания и выступание на сцену легочного дыхания наблюдается уже у Dipnoi; изменения в этом направлении наблюдаются у Polypterus, и Lepidosteus. У амфибий жаберное дыхание сохраняется прежде всего у личинок, а затем у тех Urodela, которые всю жизнь проводят и воде (Perennibranchiata в прежних системах). Жаберные щели унаследованы амфибиями от рыбообразных предков. Жаберные дуги обнаружены у стегоцефал, у личинок и у некоторых взрослых (Branchiosauridae). Все современные амфибии в личиночном состоянии дышат жабрами. Нормально у них закладывается 5 висцеральных мешков и 6-й недоразвитый. Ho наружу открываются не все: жаберных щелей образуется 4 или еще меньше. Иногда щелей значительно меньше, чем дуг. Наличие щелей и дуг является доказательством происхождения амфибий от рыб. Внутренние жабры, гомологичные жабрам рыб, встречаются, однако, только у личинок Anura в виде коротких выростов покровов на дугах, разделяющих жаберные щели. Мягкой жаберной крышечкой (operculum), разрастающейся со стороны гиоидной дуги, жаберная область прикрывается снаружи. Жаберные крышечки правой и левой сторон сливаются между собой с нижней стороны, оставляя у некоторых Anura парные отверстия, а у большинства - одно непарное на левой стороне тела.
На ранних стадиях развития у личинок Anura и у всех других амфибий имеются только наружные жабры, гомологичные, видимо, наружным жабрам личинок Polypterini и Dipnoi. У Apoda и у Anura наружные жабры существуют лишь в личиночном периоде, на ранних стадиях развития, у Urodela же, вторично вернувшихся к водной жизни, они сохраняются всю жизнь. Отсюда название для этих амфибий - постоянно жаберные (Perennibranchiata), хотя название это, как было сказано, обнимает группы амфибий, имеющих различное происхождение. Наружные жабры, вероятно унаследованы амфибиями от кистеперых рыб.
Легкие амфибии имеют вид длинных цилиндрических мешков с тонкими стенками (у Urodela) или более коротких (у Anura). У безногих правое легкое развито гораздо больше левого. Появились легкие у предков четвероногих еще задолго до выхода па сушу. Мы видим такие же легкие у двоякодышащих. Появились они, очевидно, в качестве дополнительного органа дыхания вследствие недостаточного развития жаберного дыхания, с одной стороны, и возможно неблагоприятных для дыхания условий в пересыхавших и портившихся водах - с другой. Задняя часть жаберной полости развилась у них в дополнительный дыхательный орган. Первоначально этот орган, имевший вид двулопастного мешка, открывавшегося на нижней стороне глотки, был несовершенен: стенки его должны были быть топкими, хотя и обильно снабженными кровью, со слабо развитыми или почти неразвитыми перегородками. Как все жаберные выпячивания (щели), он имел гладкую висцеральную мускулатуру и иннервировался блуждающим первом.
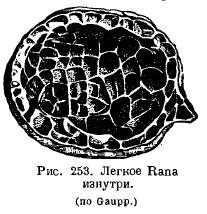
Легкие амфибий мало изменились в сравнении с этим: у водных Urodela легкие выполняют скорее роль гидростатического аппарата и имеют гладкую внутреннюю поверхность; высота организации их даже ниже, чем у Dipnoi, Нормально же у амфибий внутренняя поверхность легких является ячеистой благодаря тому, что в полость легкого вдается система перекладин (рис. 253). Весьма интересно, что чем более наземным является тот или иной вид, тем более развитыми являются перекладины в легких: у жабы легкое более ячеисто, чему лягушки. У рода Ascaphus, живущего в горных ручьях, в воде, богатой кислородом,сильно развито кожное дыхание, легкие же, наоборот, маленькие и слабо снабжаются кровью. Ряд амфибий из подотряда Salamandroidea (Salamandrina, Plethodon, Spelerpes, Batrachoseps и др.) совершенно утратил легкие, взамен которого глоточное и кожное дыхание сильно развилось. .
Легочные мешки в простейшем случае соединяются между собой впереди, открываясь непосредственно в глотку продольной щелью, поддерживаемой по сторонам хрящевыми полосками. Эти хрящевые полоски при помощи прикрепляющихся к ним мускулов могут расширять и суживать гортанную щель.
Хрящи эти происходят из последней жаберной дуги и в простейшем виде встречаются у некоторых Urodela. От этих хрящей могут обособляться хрящики, называемые перстневидными хрящами. Их можно сравнивать с черпаловидными хрящами (cartilagines arythenoidea) высших позвоночных. Некоторые Urodela, а также Apoda имеют довольно длинное дыхательное горло, поддерживаемое хрящевыми кольцами. У Anura слизистая оболочка в гортани образует голосовые связки. Гортань имеет сложную мускулатуру. На дне или по углам рта находятся резонаторы, надувающиеся при квакании.
Механизм дыхания наземных амфибий имеет своим корнем рефлексы, наблюдающиеся у рыб и водных амфибий. Ближе всего к дыханию рыб дыхание личинок Anura, имеющих внутренние жабры, оперкулярную складку и образованную их срастанием жаберную полость, открывающуюся наружу одним отверстием. Кроме того, у личинок амфибий ротовая полость обильно снабжена кровью. Забирая воду в рот и проталкивая ее путем поднятия челюстей через ноздри, личинки увеличивают газовый обмен в ротовой полости. Когда же личинки подрастают, они поднимаются к поверхности, где заглатывают воздух подобно цератоду, и поднятием дна ротоглоточной полости проталкивают воздух в легкие. У водных Urodela наблюдается сходный акт. При опускании дна ротоглоточной полости и при закрытых сзади жаберных отверстиях вода насасывается в ротовую полость через рот или ноздри или через то и другое вместе. Путем последующего поднятия дна ротовой полости при закрытых ноздрях вода выталкивается через жаберные щели. Благодаря этим движениям слизистая оболочка рта и глотки вступает в соприкосновение с новыми массами воды, а у жабер получается движение, обновляющее дыхательную среду.
У наземных амфибий механизм дыхания является актом заглатывания воздуха благодаря опусканию мускулистого дна ротовой полости и проталкиванию его в легкие вследствие поднятия дна. Таким образом, дыхание наземных амфибий является актом, осуществляемым по типу нагнетательного насоса, преобладающего у низших рыб. Непосредственной основой, на которой он развивается, является механизм дыхания постоянножаберных амфибий. Этот последний, наблюдаемый, например, у Necturus, должен был развиться у отдаленных рыбообразных предков амфибий. Из него уже развился более сложный тип дыхания наземных - Anura.
У безлегочных саламандр сильно развит газообмен внутриротовой и глоточной полости, совершающийся при помощи частых, до 120-170 колебаний в минуту диафрагмы рта (у лягушек их 30).
Вообще следует сказать, что легочное дыхание у амфибий в целом является вспомогательным способом дыхания. В этом также имеется указание и па его филогенетическое происхождение.
Дыхание современных амфибий ни в коем случае не могло быть источником развития дыхания высших Tetrapoda (дыхания при помощи поднятия ребер, расширения грудной клетки и насасывания таким образом воздуха). Последний тип мог возникнуть, во всяком случае наметиться у древнейших вымерших амфибий, имевших длинные ребра.
Уменьшение числа жабр.
Увеличение дыхательной поверхности за счет образования жаберных лепестков.
Образование жаберных капилляров.
У ланцетника боковые стенки глотки пронизаны многочисленными (до 150 пар) косо расположенными жаберными щелями. К межжаберным перегородкам подходят приносящие жаберные артерии, а отходят выносящие жаберные артерии. Когда вода омывает межжаберные перегородки, происходит газообмен между проходящей водой и кровью, которая течет по тонким сосудам перегородок. Жаберные артерии на капилляры не разветвляются. Кроме того, кислород проступает в организм животного через капилляры кожи.
У первичноводных позвоночных животных (бесчелюстные и рыбы), как и у низших хордовых, образуются жаберные щели, соединяющие полость глотки с внешней средой. У круглоротых, из выстилающей жаберные щели энтодермы, формируются жаберные мешки (у рыб жабры развиваются из эктодермы). Внутренняя поверхность мешков покрыта многочисленными складками – жаберными лепестками, в стенках которых ветвится густая сеть капилляров. Мешок внутренним узким каналом открывается в глотку (у взрослых миног – в дыхательную трубку), а наружным – на боковой поверхности тела животного. У миксин от 5 до 16 пар жаберных мешков, у семейства бделлостомовые каждый из них открывается наружу самостоятельным отверстием, а у семейства миксиновые все наружные жаберные ходы на каждой стороне сливаются в один канал, который открывается наружу одним отверстием, расположенным далеко сзади. У миног 7 пар жаберных мешков, каждый из которых открывается наружу самостоятельным отверстием. Дыхание осуществляется путем ритмичных сжатий и расслаблений мышечной стенки жаберной области. У непитающихся миног вода поступает в дыхательную трубку из ротовой полости, затем омывает лепестки жаберных мешков, обеспечивая газообмен, и удаляется через наружные жаберные ходы. У питающихся круглоротых вода поступает и выводится через наружные отверстия жаберных мешков.
Дыхательная система рыб имеет специализированные органы газообмена – эктодермальные жабры, которые либо расположены на межжаберных перегородках, как у хрящевых рыб, либо непосредственно отходят от жаберных дуг, как у костных рыб. Обмен газов в жабрах позвоночных построен по типу «противоточных систем»: при встречном движении кровь контактирует с богатой кислородом водой, что обеспечивает ее эффективное насыщение. Увеличение поверхности поглощения кислорода благодаря образованию жабр сопровождалось уменьшением числа жаберных щелей у позвоночных по сравнению с низшими хордовыми. У цельноголовых (из хрящевых рыб) намечается редукция межжаберных перегородок и образуется кожистая жаберная крышка, прикрывающая снаружи жабры. У костных рыб в жаберной крышке появляется костный скелет, а межжаберные перегородки редуцируются, что способствует более интенсивному омыванию водой жаберных лепестков. Наряду с газообменом жабры рыб участвуют в водном и солевом обмене, в выведении из организма аммиака и мочевины. В качестве добавочных органов дыхания у отдельных групп рыб функционируют кожа, плавательный пузырь, надглоточные лабиринты и специализированные участки кишечной трубки. У двоякодышащих и многоперообразных рыб появляются органы воздушного дыхания – легкие. Легкие возникают как парные выросты брюшной части глотки в области последней жаберной щели и коротким каналом связаны с пищеводом. Стенки этого выроста тонкие и обильно снабжаются кровью.
Направления эволюции легочного типа дыхания
Появление и дифференцировка дыхательных путей.
Дифференцировка легкого и увеличение дыхательной поверхности.
Развитие вспомогательных органов (грудной клетки).
У земноводных в поглощении кислорода и выделении углекислоты участвуют: у личинок – кожа, наружные и внутренние жабры, у взрослых – легкие, кожа и слизистая оболочка ротоглоточной полости. У некоторых видов хвостатых земноводных (сирен, протеев) и у взрослых сохраняются жабры и недоразвиваются или редуцируются легкие. Соотношение легочного и другого типов газообмена неодинаково: у видов влажных местообитаний в газообмене доминирует кожное дыхание, у обитателей сухих мест большая часть кислорода поступает через легкие, но кожа играет существенную роль в выделении углекислого газа. Дыхательная система взрослых амфибий включает ротоглоточную, гортанно-трахейную полости и мешковидные легкие, стенки которых оплетены густой сетью капилляров. У бесхвостых земноводных имеется общая гортанно-трахейная камера, у хвостатых она разделяется на гортань и трахею. В гортани появляются черпаловидные хрящи, которые поддерживают ее стенку и голосовые связки. Легкие хвостатых амфибий представляют собой два тонкостенных мешка, не имеющих перегородок. У бесхвостых внутри легочных мешков на стенках имеются перегородки, которые увеличивают поверхность газообмена (легкие ячеистые). У земноводных нет ребер, и акт дыхания происходит путем нагнетания воздуха при вдохе (за счет увеличения, а затем уменьшения объема ротоглоточной полости) и выталкивания воздуха при выдохе (за счет эластичности стенок легких и брюшной мускулатуры).
У рептилий отмечается дальнейшая дифференцировка дыхательных путей и значительное увеличение функциональной поверхности газообмена в легких. Воздухоносные пути подразделяются на носовую полость (она объединена с ротовой полостью, но у крокодилов и черепах эти полости разделены костным небом), гортань, трахею и два бронха. Стенки гортани поддерживают парные черпаловидные и непарный перстневидный хрящи. У ящериц и змей внутренние стенки легочных мешков имеют складчатое ячеистое строение. У черепах и крокодилов сложная система перегородок вдается во внутреннюю полость легкого настолько глубоко, что легкое приобретает губчатое строение. Формируется грудная клетка: ребра подвижно соединены с позвоночником и грудиной, развиваются межреберные мышцы. Акт дыхания осуществляется благодаря изменению объема грудной клетки (реберный тип дыхания). У черепах сохраняется ротоглоточный тип нагнетания воздуха. У водных черепах в воде дополнительными органами дыхания служат богатые капиллярами выросты глотки и клоаки (анальные пузыри). Кожное дыхание у рептилий отсутствует.
У птиц воздухоносные пути представлены носовой полостью, гортанью, которая поддерживается черпаловидными и перстневидным хрящами, длинной трахеей и системой бронхов. Легкие небольшие, плотные и мало растяжимые приращены к ребрам по бокам позвоночного столба. Первичные бронхи образуются при разделении нижней части трахеи и входят в ткань соответствующего легкого, где распадаются на 15–20 вторичных бронхов, большинство которых заканчиваются слепо, а часть сообщается с воздушными мешками. Вторичные бронхи связаны между собой более мелкими парабронхами, от которых отходит множество тонкостенных ячеистых бронхиол. Оплетенные кровеносными сосудами бронхиолы образуют морфофункциональную структуру легкого. С легкими птиц связаны воздушные мешки – прозрачные эластичные тонкостенные выросты слизистой оболочки вторичных бронхов. Объем воздушных мешков примерно в 10 раз превышает объем легких. Они играют очень важную роль в осуществлении своеобразного дыхательного акта птиц: в легкие как при вдохе, так и при выдохе поступает воздух с высоким содержанием кислорода – «двойное дыхание». Помимо интенсификации дыхания воздушные мешки предотвращают перегрев организма при интенсивном движении. Повышение внутрибрюшного давления при выдохе способствует дефекации. Ныряющие птицы, увеличивая давление в воздушных мешках, могут уменьшать объем и тем самым увеличивать плотность, что облегчает погружение в воду. Кожное дыхание у птиц отсутствует.
У млекопитающих наблюдается дальнейшая дифференцировка дыхательных путей. Образуется носовая полость, носоглотка, вход в гортань прикрывает надгортанник (у всех наземных позвоночных кроме млекопитающих гортанная щель замыкается специальной мускулатурой), в гортани появляется щитовидный хрящ, далее идет трахея, которая разветвляется на два бронха, идущие в правое и левое легкое. В легких бронхи многократно ветвятся и заканчиваются бронхиолами и альвеолами (число альвеол от 6 до 500 млн.), это значительно увеличивает дыхательную поверхность. Газообмен происходит в альвеолярных ходах и альвеолах, стенки которых густо оплетены кровеносными сосудами. Морфофункциональной единицей легкого млекопитающих является легочной ацинус, который образуется в результате ветвления терминальной бронхиолы. Формируется грудная клетка, которая отделяется диафрагмой от брюшной полости. Число дыхательных движений от 8 до 200. Дыхательные движения осуществляются двумя способами: за счет изменения объема грудной клетки (реберное дыхание) и за счет деятельности диафрагмальной мышцы (диафрагмальное дыхание). У высших млекопитающих развито кожное дыхание через систему кожных капилляров, которое играет большую роль в газообмене.
